Die ersten Tiere
Die primitivsten Lebewesen sind einzellig: Der gesamte Organismus besteht aus einer einzigen Zelle. Einige Organismen können als Einzelzellen oder als Gruppen von Zellen in einer Kolonie leben. Wenn die Zellen einer Kolonie getrennt werden, können sie als Einzelzellen weiterleben oder eine neue Kolonie bilden. Keine Zelle ist auf eine bestimmte Aufgabe spezialisiert. Die moderne Alge Volvox ist ein gutes Beispiel dafür: Das nebenstehende Foto zeigt eine Volvox-Kolonie.
Mehrzelligkeit hat sich unabhängig voneinander ein paar Dutzend Mal in Pflanzen, Rotalgen, Braunalgen, Pilzen und Tieren entwickelt. Zu den Rotalgen in der Gruppe der Volvocales gehören zum Beispiel einzellige Arten (Chlamydomonas), generalisierte vierzellige Arten (Basichlamys) und Arten, die Kolonien bilden, die 50.000 Zellen umfassen können (Volvox) (Sachs, 2008).
Während viele Algen in diesen einfachen Kolonien leben, haben einige Algen tatsächlich Zellen, die sich auf verschiedene Aufgaben spezialisiert haben (sie haben Gewebe, die in ihrer Funktion wurzelähnlich und in ihrer Funktion blattähnlich sind).
Die ersten Fossilien von Algen sind aus der Zeit vor etwa 2,1 Milliarden Jahren bekannt und ab etwa 1 Milliarde Jahren nimmt die Vielfalt der bekannten Algenarten zu. Eukaryotische Algen scheinen sich vor etwa 600 Millionen Jahren, nach den Eiszeiten des Endproterozoikums, plötzlich diversifiziert zu haben (Cavalier-Smith, 2000).
Vor 570 Millionen Jahren sind Algen mit einer Vielzahl von anatomischen und reproduktiven Merkmalen bekannt (Xiao, 1998). Der Fossilbericht hat eine Vielfalt an prähistorischen Algen wie die unten abgebildeten erhalten.
FUNGI
Viele der Fossilien aus dem Prä-Ediacaran repräsentieren einfache mehrzellige Kolonien von Bakterien, eukaryotische Algen (einschließlich Grün- und Rotalgen) und Formen, die Ähnlichkeiten mit Pilzen aufweisen. (Butterfield, 2009; Xiao, 2009).
Karnivore Pilze, die Hyphenringe bilden, um Nematoden zu erbeuten, wurden in 100 Millionen Jahre altem Bernstein beobachtet (Schmidt, 2007).
DIE ERSTEN TIERE
Die Ediacaran-Fauna ist eine Ansammlung von Organismen, die nach den Ediacara-Hügeln in Australien benannt ist, wo sie zuerst gefunden wurden. Fossilien aus dem Ediacaran sind aus mehr als 30 Ländern und allen Kontinenten außer der Antarktis und Südamerika bekannt (De, 2005; McCall, 2006).Mehrere tausend Exemplare, die mehr als 270 Arten umfassen, wurden aus der Ediacaran-Fauna beschrieben und viele müssen noch beschrieben werden (Fedonkin, 1990; Minnerop, 2000; Waggoner, 1998; Shen, 2008). Das erste Tier aus dem Ediacaran wurde 1872 beschrieben, gefolgt von einem zweiten Fund in den Jahren 1908-14 (McCall, 2006).
- Sie sind alt
Fossile Schichten, die auf ein Alter von mehr als 1,2 Milliarden Jahren datiert wurden, enthalten Tiere – nicht nur Fossilien von strahlenförmigen Weichkörperorganismen, sondern auch Höhlen, die von schleimproduzierenden Würmern angelegt worden zu sein scheinen (Rasmussen, 2002; McCall, 2006). Es gibt eine Fundstelle in Indien, deren Wurmhöhlen auf 1,1 Milliarden Jahre datiert wurden, obwohl einige dieses Datum bestreiten. Kürzlich wurde eine andere Fundstelle mit offensichtlichen Anzeichen von primitiven Tieren (Quallenverwandte) und Wurmgruben auf ein Alter von höchstens 1,2 Milliarden Jahren datiert (Seilacher, 1998; Kerr, 1998b; Morris, 1993). Andere Ediacaran-Fossilien sind vor 610-600 Mio. Jahren bekannt und vielfältigere Organismengruppen vor 555,3 Mio. Jahren (McMenamin, 1996, Minnerop, 2000).Präkambrische Gesteinsschichten, die 580 Mio. Jahre alt sind, besitzen Fossilien von kleinen Bilateralen; größere Bilaterale sind ab einem Datum vor 555 Mio. Jahren vorhanden (Condon, 2005).Sie wurden weitgehend durch die Tiere der „kambrischen Explosion“ ersetzt, die vor etwa 545 Mio. Jahren begann. Die Ediacaran-Periode ist definiert als die Zeit vor 635 Millionen Jahren nach der Marinoan Snowball Earth-Vereisung (obwohl die Gaskiers-Vereisung vor 562 Millionen Jahren innerhalb der Ediacaran-Periode stattfand) (Xiao, 2009).
Molekulare Vergleiche moderner Organismen stützen ebenfalls die Schlussfolgerung, dass Tiere bereits vor dem Kambrium eine Vielzahl von Linien hervorgebracht haben. So legt eine Studie von 129 Proteinen nahe, dass Eukaryoten vor 950 bis 1.260 Millionen Jahren entstanden sind, Tiere vor 761 bis 957 Millionen Jahren, und Coelomaten-Linien trennten sich vor 642 bis 761 Millionen Jahren (Douzery, 2004). Andere molekulare Schätzungen für den Ursprung der Eumetazoen (Nesseltiere und Bilaterale), die von Studien zur molekularen Uhr geliefert wurden, variieren von vor 570 Millionen Jahren bis vor 1.100 Millionen Jahren. Der Ursprung der Bilateralen Tiere wurde auf 580 Millionen Jahre geschätzt (Peterson, 2005). Fossilien von Tieren, die in Australien, Indien und Texas gefunden wurden, wurden auf ein Alter von mehr als 1 Milliarde Jahre datiert und ein segmentiertes Tier aus Russland wurde auf ein Alter von 890 Millionen bis 1 Milliarde Jahre datiert (De, 2005). Aufgrund der aufregenden Entdeckungen von präkambrischen Tieren wurde der geologischen Zeitskala eine neue Periode hinzugefügt, was das erste Mal ist, dass eine neue Periode seit 1891 (als das Karbon geteilt wurde) stratigraphisch definiert wurde. Die Ediacaran-Periode ist definiert als die Zeit zwischen 630 Millionen Jahren vor heute und dem Beginn des Kambriums, vor etwa 542 Millionen Jahren (Knoll, 2004).
Wie aussagekräftig sind diese Daten? Bis vor kurzem dachte man, dass eine Vielfalt komplexer Tiere plötzlich in der „kambrischen Explosion“ vor etwa 545 Millionen Jahren auftauchte. Heute wissen wir, dass Tiere schon mindestens 60 Millionen Jahre vorher existierten und vielleicht sogar 600 Millionen Jahre vor dem Kambrium. Sechzig Millionen Jahre mögen angesichts der riesigen Zeitspanne im Präkambrium nicht wie eine so lange Zeit erscheinen, aber in einem solchen Zeitraum kann eine große Menge an tierischer Evolution stattfinden. Es dauerte etwa sechzig Millionen Jahre, bis sich aus den rhipidistischen Fischen die ersten Amphibien entwickelten und etwa 60 Millionen Jahre für die anschließende Diversifizierung dieser Amphibien in eine Vielzahl von Linien, einschließlich der ersten Reptilien. Ein Zeitraum von sechzig Millionen Jahren, der am Ende des Perms beginnt, würde zwei Massenaussterben beinhalten. Es beginnt mit einer Welt, die von synapsiden Reptilien und primitiven Archosauriern dominiert wird und endet im Jura mit seinen verschiedenen Linien von Dinosauriern, Flugsauriern und Meeresreptilien. In etwa sechzig Millionen Jahren strahlten die primitiven Säugetiere, die das Aussterben der Dinosaurier überlebten, in die vielfältigen fossilen und lebenden Gruppen des Känozoikums aus.


2) SIE SIND EINFACHE TIERE
Die meisten Organismen aus dem Ediacaran sind unter einem Zentimeter groß, aber einige sind mehrere Meter lang. Sie sind seltsam und die meisten unterscheiden sich stark von allem, was heute lebt (viele bezeichnen sie als die Gruppe Vendobionta). Praktisch alle haben keinen Kopf oder Schwanz, keine Innen- oder Außenseite, keine Vorder- oder Rückseite, keine Zähne oder Augen. Die meisten haben keine offensichtlichen Kreislauf-, Nerven- oder Verdauungssysteme oder innere Organe jeglicher Art. Die meisten sind sehr dünn und flach – ein Zustand, der für Tiere ohne Kreislauf-, Atmungs-, Verdauungs- oder Ausscheidungssysteme unerlässlich ist. Einige Fundorte weisen eine Vielfalt an Tieren auf, mit bis zu 30 verschiedenen Taxa (Buss, 1994).
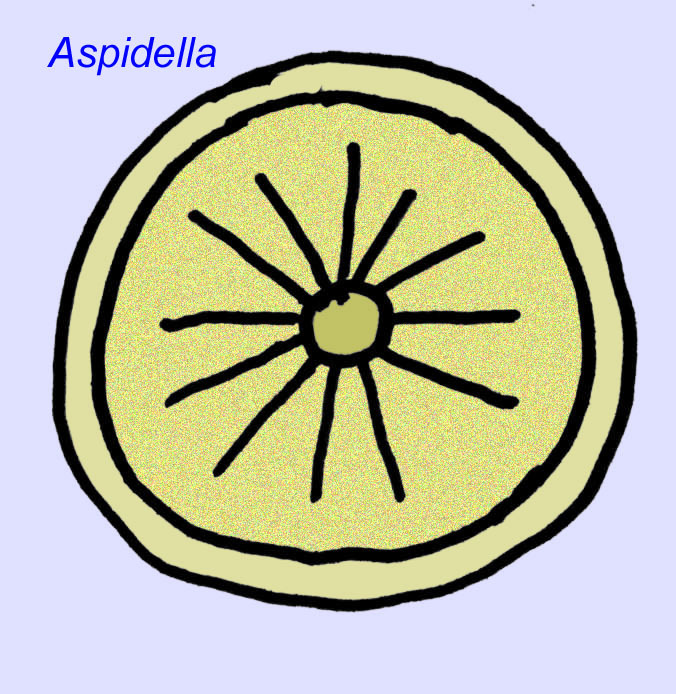
Was sind das für Tiere? In der Vergangenheit gab es eine Vielzahl von Meinungen über diese Organismen: Einige klassifizierten sie als Pflanzen (die Schlussfolgerung ab dem Ersten Weltkrieg), sehr große Einzeller, einzigartige Flechten, eine völlig ausgestorbene Gruppe von Tieren und eine Gruppe von Tieren, die die Vorfahren der modernen Tiere umfasst. Ursprünglich dachte man, dass sie lange vor dem Aufkommen der modernen Tiergruppen ausgestorben sind, jetzt ist es offensichtlich, dass einige Gruppen bis ins Kambrium überlebt haben und möglicherweise Fossilien von nicht-metazoischen Tieren darstellen (Droser, 2006). Die Individuendichte des Scheibentieres Aspidella deutet darauf hin, dass die Biomasse der bodenbewohnenden Tiere im Präkambrium annähernd so groß gewesen sein könnte wie in modernen Lebensgemeinschaften (Gehling, 2000).
Viele der prä-Ediacaran-Fossilien stellen einfache mehrzellige Kolonien von Bakterien, eukaryotischen Algen (einschließlich Grün- und Rotalgen) und Formen dar, die Ähnlichkeiten mit Pilzen aufweisen. Einige Fossilien aus dem Neoproterozoikum wurden als Schwämme und Larven von Schwämmen interpretiert (Butterfield, 2009).
Schwämme, die zu den einfachsten Tieren gehören, wurden in Sedimenten aus dem Ediacaran identifiziert (z. B. die Gattung Palaeophragmodictya), die einigen paläozoischen Schwämmen ähnlich sind. Diese Schwämme konnten 1,5-10 cm im Durchmesser messen (Gehling, 1996; Kerr, 1998a). Schwammnadeln sind in Ediacaran-Gesteinen vorhanden (Xiao, 2000).
Einige Ediacaran-Fossilien wurden als Pilze und basale Tiere interpretiert, die primitiver als Schwämme sind (Xiao, 2009).
Nesseltiere
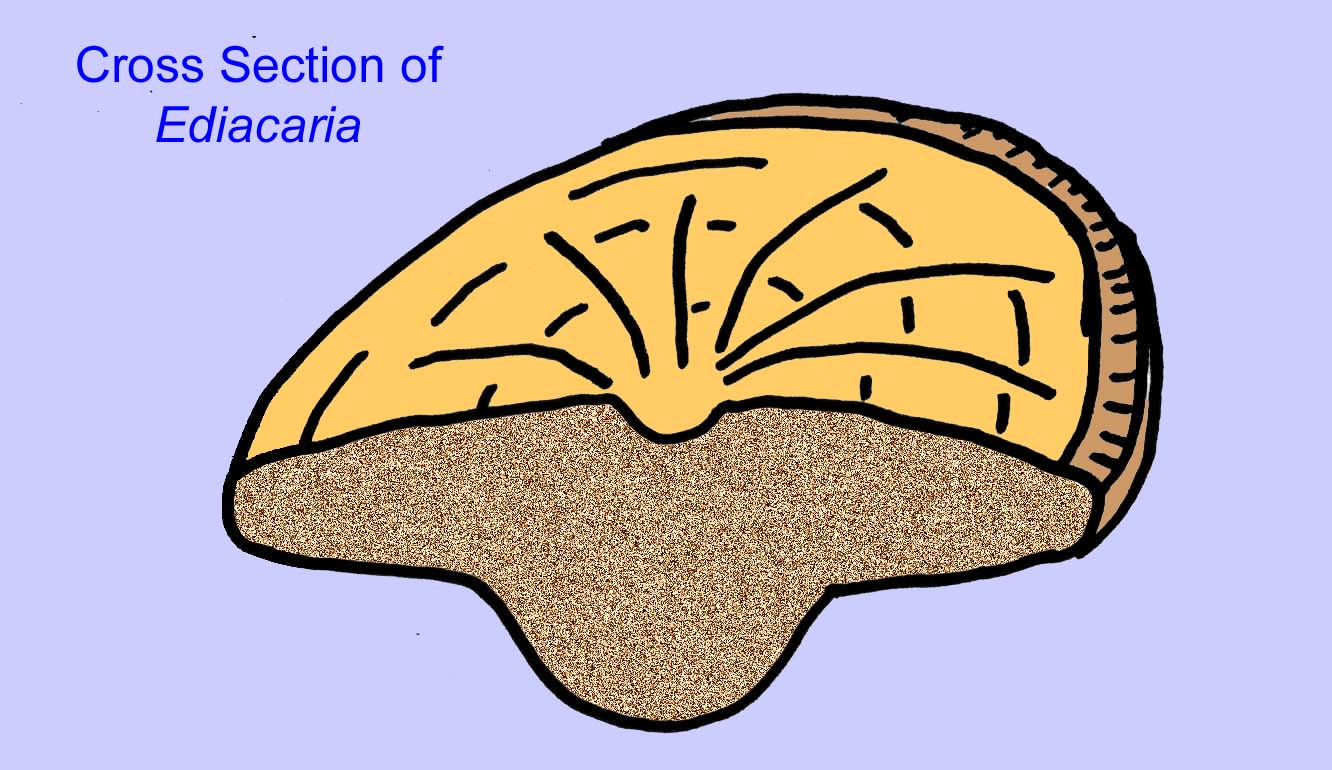
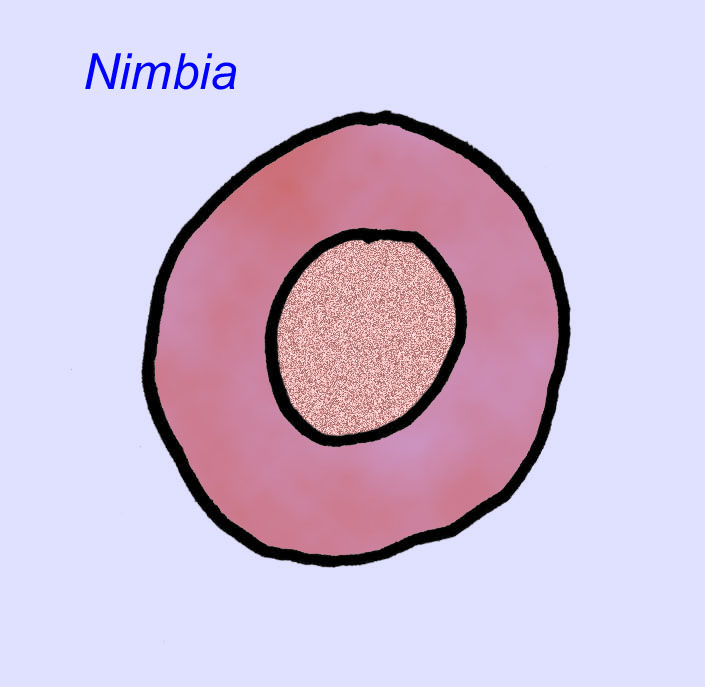
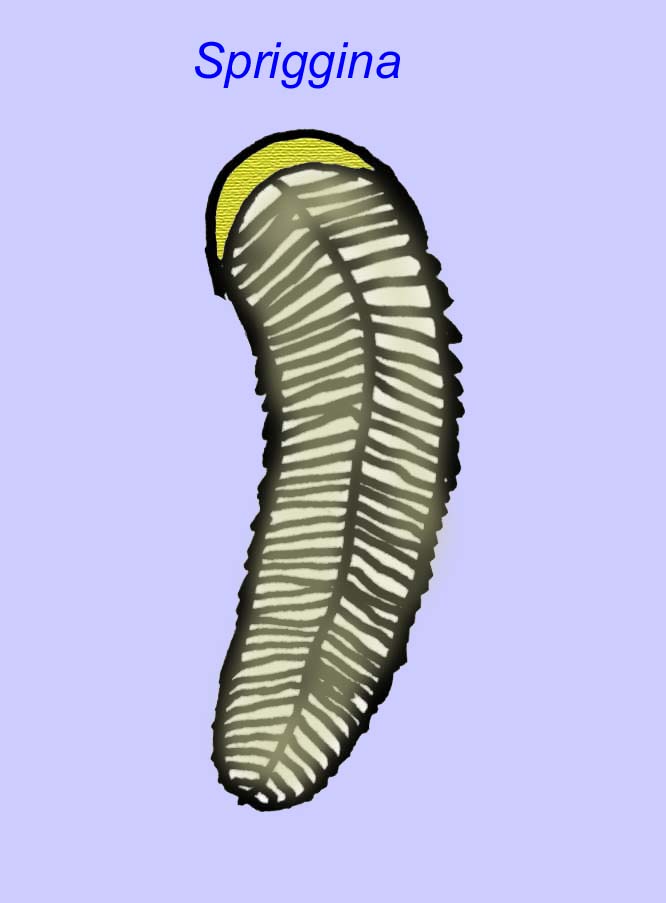
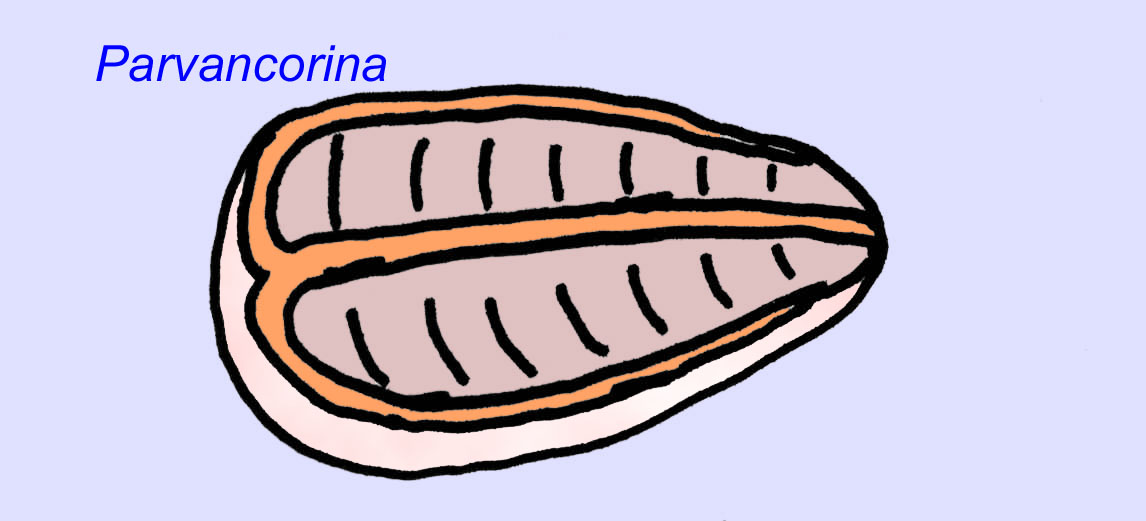
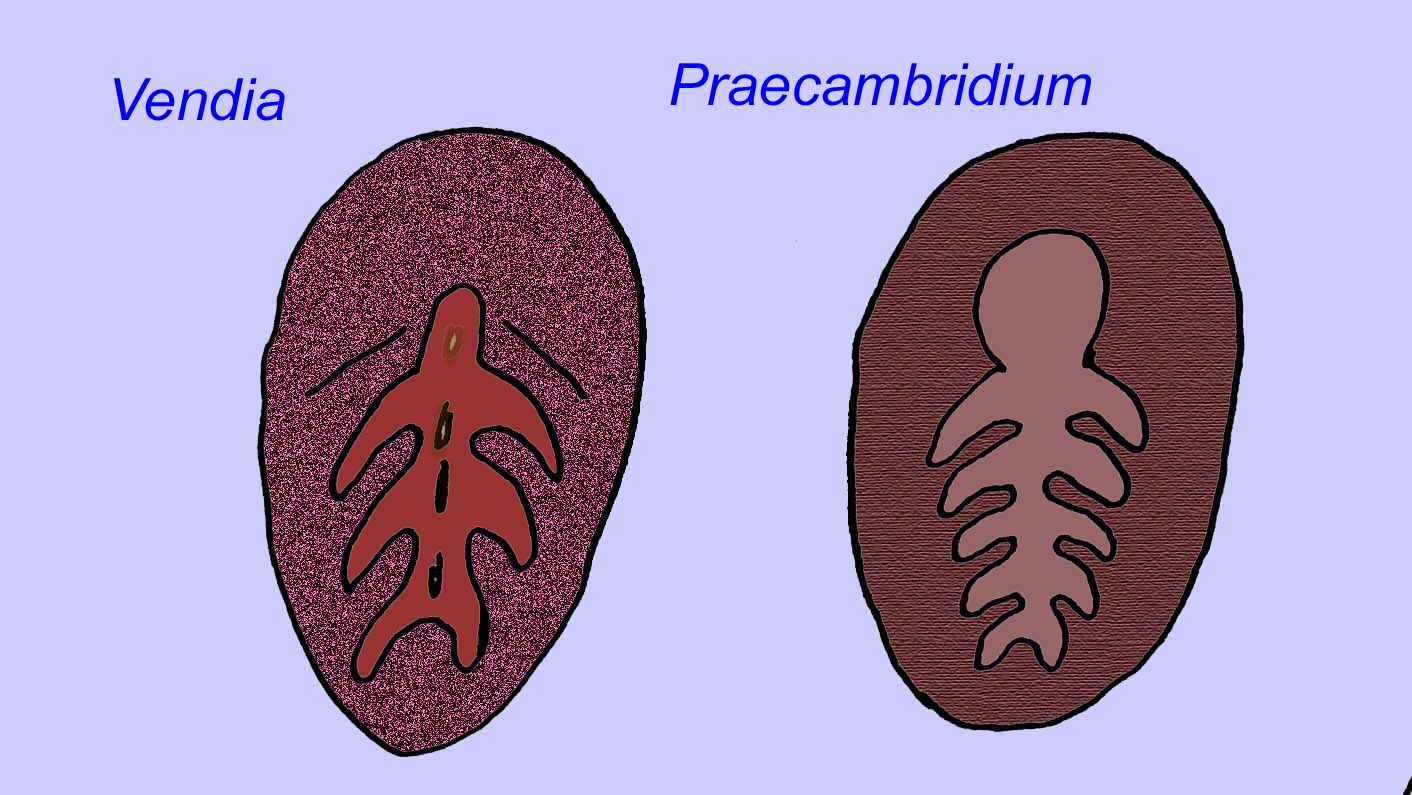
Es besteht ein allgemeiner Konsens, dass viele Ediacaran-Fossilien als frühe Nesseltiere klassifiziert werden sollten, eine Gruppe von Tieren, zu denen auch die modernen Quallen, Seepferdchen und Seefächer gehören. Einige Formen (wie z. B. Ediacaria und Nimbia) sind radialsymmetrisch und ähneln oberflächlich den Nesseltieren, aber es fehlen jegliche Anzeichen eines Mundes, was ein Grund sein kann, sie aus dieser Gruppe auszuschließen. (Es ist nicht sicher, ob die ersten Nesseltiere einen Mund hatten, wie es bei allen späteren Formen der Fall ist.) Andere, wie z. B. Albumares brunsae, besitzen ein gastrovaskuläres System, das dem der Nesseltiere ähnelt (Lipps, 1992; Sokolov, 1985). Viele haben eine zentrale scheibenförmige Struktur auf einer Seite ihres flachen, runden Körpers (Crimes, 1995; Bertrand-Sarfati, 1995). Einige Ediacaran-Tiere (Spriggina, Parvancorina und Praecambridium) haben Unterschiede zwischen ihrer Ober- und Unterseite (posterior und anterior). Einige, wie z. B. Ediacaria, hatten radiale Linien und periphere Verdickungen (Crimes, 1995). Einige der Ediacaran-Nesseltiere wurden als eine separate, aber ausgestorbene Unterklasse klassifiziert (McCall, 2006).
Während diesen Tieren ein mineralisiertes Skelett fehlte, besaßen die meisten oder sogar alle (wie Cyclomedusa und Ernietta) eine Art von starrem Skelett, das in der Lage war, einen Abdruck im Schlamm zu hinterlassen. Die Art des Gewebes, aus dem das innere Skelett bestand, ist nicht bekannt (Crimes, 1995; Dzik, 1999).
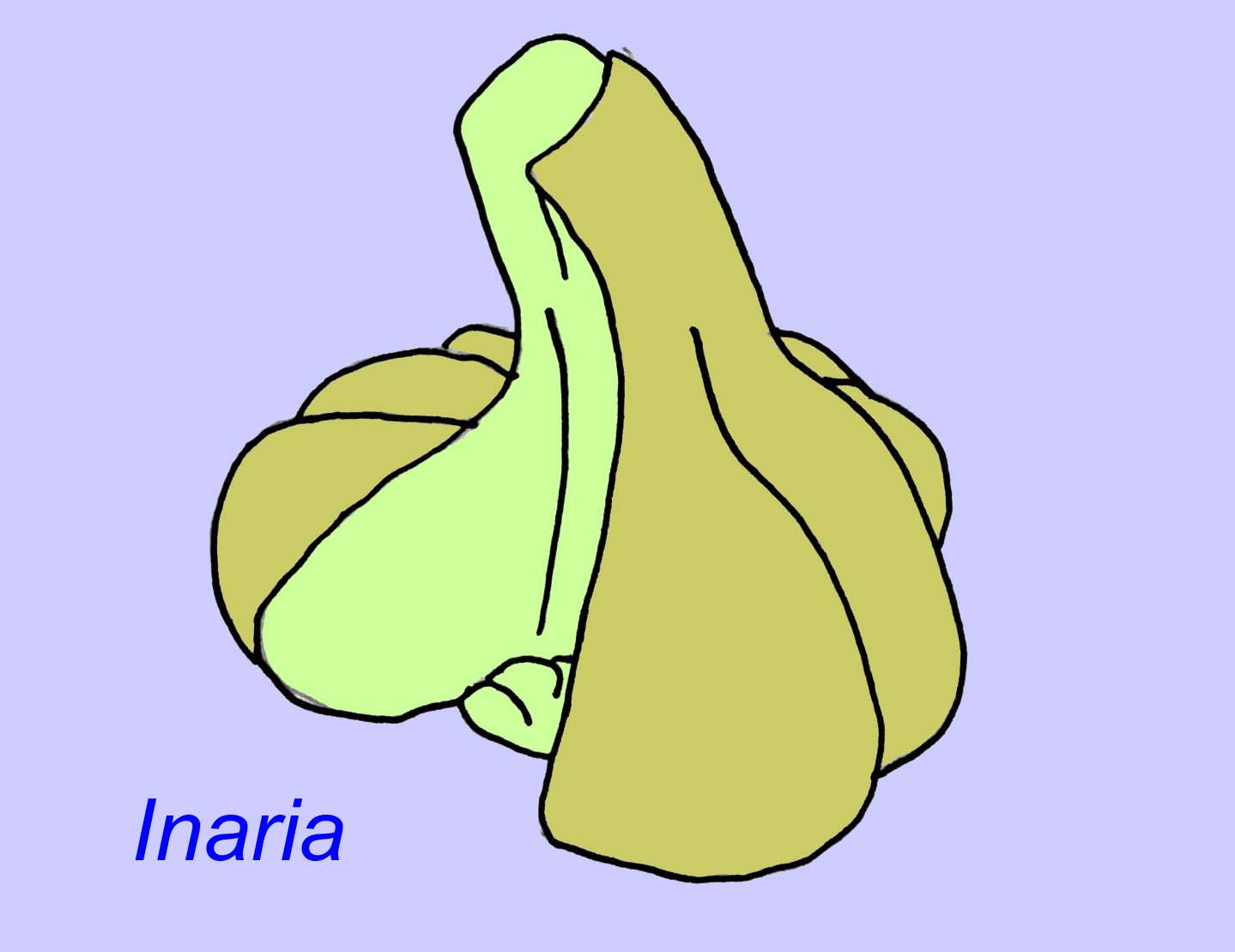
Mackenzia scheint auch ein Nesseltier zu sein (selbst wenn nicht, ist es mit ziemlicher Sicherheit ein metazoisches Tier). Es gibt indirekte Hinweise auf Nahrungsballen in einer Verdauungshöhle und auf Muskulatur (Morris, 1993a). Sinocyclocyclicus könnte ein basales Nesseltier (oder sogar ein basales Metazoon) sein, dessen apikale Knospung der einiger Nesseltiere (wie z. B. der Tabelenkorallen) ähnelt (Xiao, 2000). Inaria war ein Nesseltier der Klasse Anthozoa mit einem Mundrand, einem Pharynx und einer Kammer, die möglicherweise photosynthetisierende Organismen beherbergte, wie man sie in vielen modernen Korallen findet (McMenaman, 1990, Gehling, 1988).
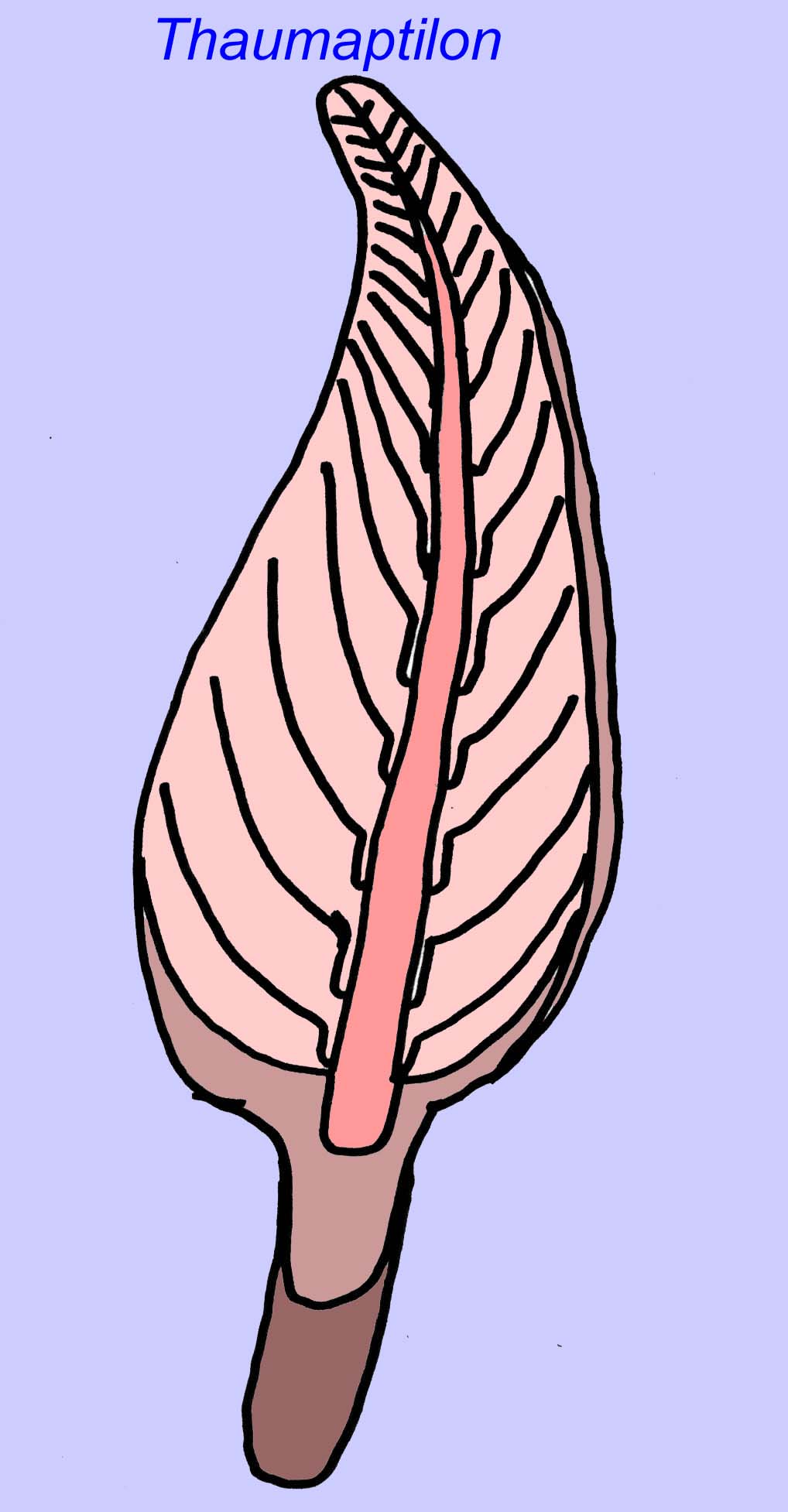
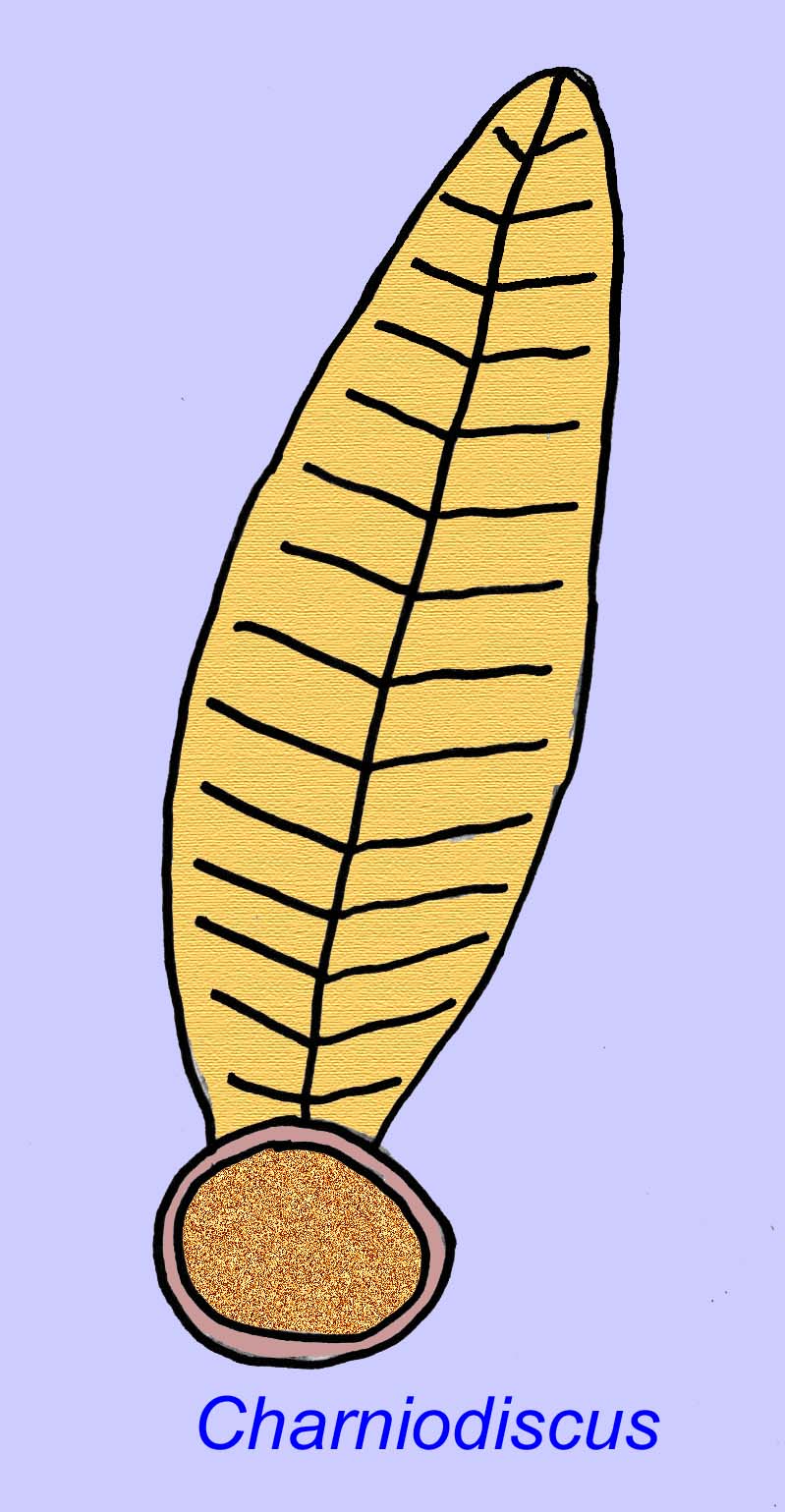
Thaumaptilon war ein frondähnliches Tier, das mit modernen Seepferdchen verwandt zu sein scheint. Es ernährte sich von partikulären Ablagerungen und benutzte eine Haltevorrichtung, um sich am Boden zu sichern (Morris, 1993a; 1993b). Der Wedel von Charniodiscus konnte eine Länge von 1,2 Metern erreichen (Sokolov, 1990).
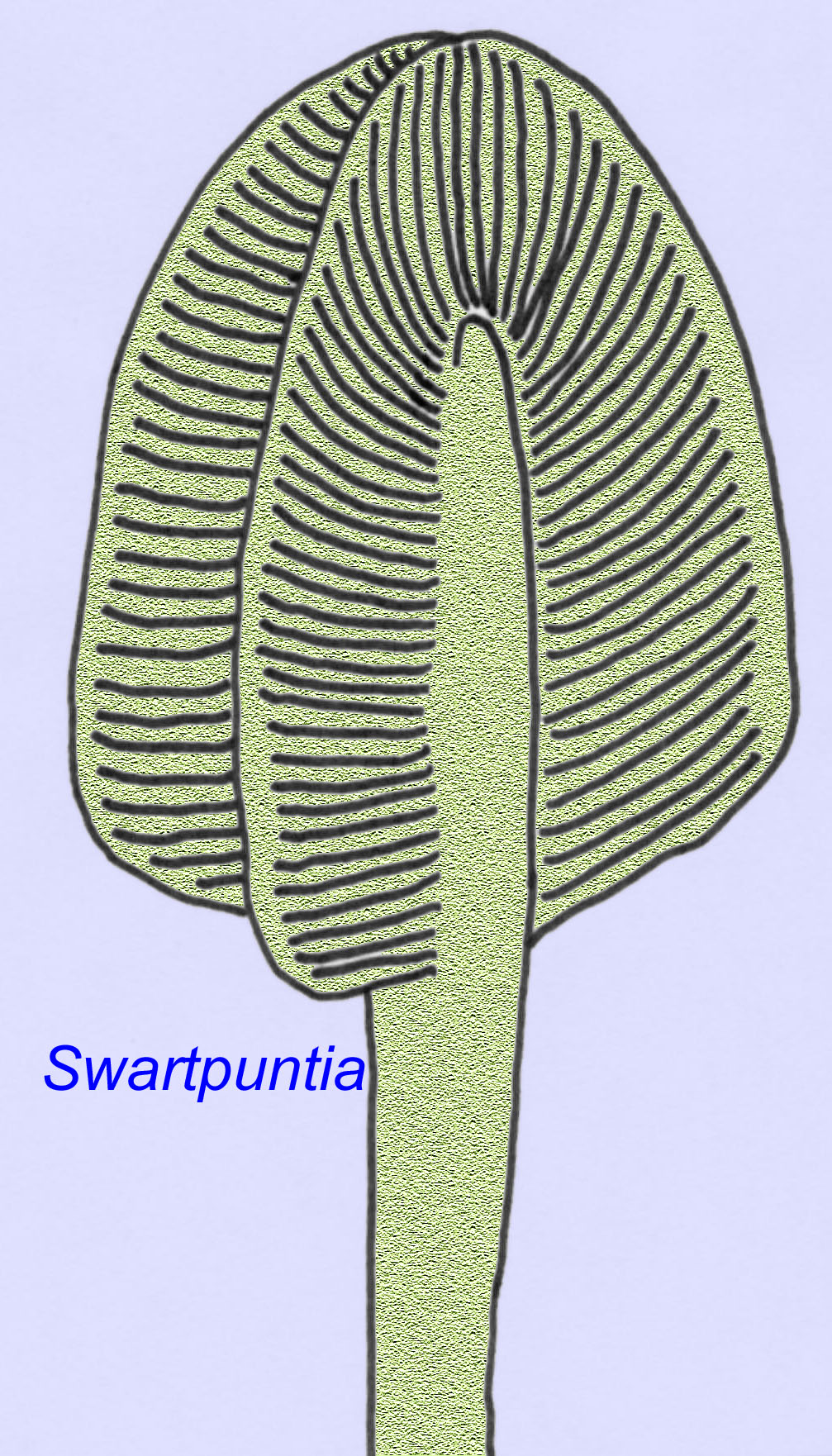
Viele Ediacaran-Tiere (z. B. Pteridinium, Swartpuntia, Charnia und Charniodiscus) besitzen einen aufrechten Stiel, der mit segmentierten Petaloiden verbunden ist.
Ein Exemplar von Charnodiscus maß 1,2 Meter in der Länge.
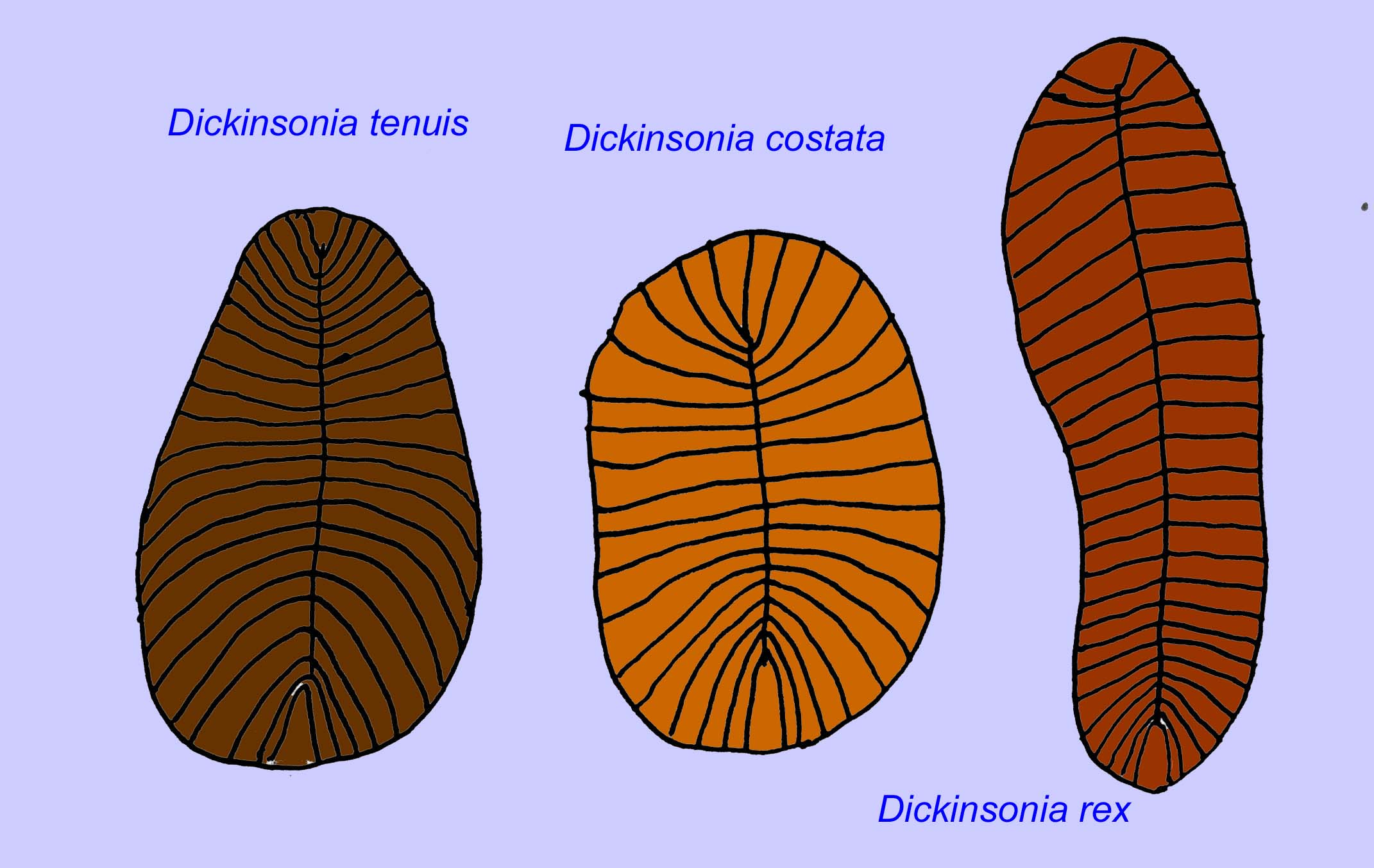
Die mikroskopische Anatomie von Swartpuntia ist der von Dickinsonia, einem frühen Wurm, sehr ähnlich (Narbonne, 1997; McCall, 2006). Aus der Zeit vor 570 Millionen Jahren sind versteinerte Embryonen bekannt, die anscheinend zu bilateralen Tieren gehören (Xiao, 1998).

Würmer aus dem Ediacaran sind aus Asien bekannt, Europa, Afrika, Australien und Nordamerika bekannt (McCall, 2006). Eine Reihe von Ediacaran-Tieren waren röhrenförmig, wie z.B. Archaeichnium, Cloudina, Corumbella und Onuphionella. Während Onuphionella mit Ringelwürmern verglichen wurde, wurde Corumbella mit den Röhren verglichen, die von den Polypen einiger Nesseltiere gebildet werden. Während Archaeichnium flexibel war, war Cloudina von einem mineralisierten Skelett bedeckt. Einige Röhren von Cloudina (maximale Länge unter 4 cm) waren ringförmig, andere waren glatt, obwohl dies Unterschiede in der Erhaltung widerspiegeln könnte (Hagadorn, 2000). Cloudina ist das erste bekannte Tier mit einem mineralisierten Skelett und 3 % der Exemplare haben Löcher in sich gebohrt, was auf Raubtiere hindeutet. Es könnte sein, dass diese Prädation die härteren Strukturen, die für so viele Tiere des Kambriums typisch sind, gefördert hat (Bengston, 1992). Cloudina ist aus dem Präkambrium Südamerikas, Europas, des Nahen Ostens und Südostasiens bekannt und Verwandte sind aus dem frühen Kambrium (vor der Verbreitung der Trilobiten) in Nordamerika bekannt (Grant, 1990). Fossilien von Cloudina sind aus einer Zeit von mindestens 6 Millionen Jahren vor dem Beginn des Kambriums bekannt (Condon, 2005).


Zusätzlich zu den beiden anerkannten Arten von Cloudina besaßen eine Reihe weiterer fossiler Arten im späten Präkambrium verkalkte Schalen (McCall, 2006). Cloudina, Namacalanthus und Conotubus sind skelettierte präkambrische Tiere, die auch im frühen Kambrium bekannt sind (Shu, 2008).
Eine Reihe von Würmern wie Paleolina evenkiana und Planolites beverlyensis wurden am Ende des Proterozoikums und bis ins Kambrium gefunden. Planolites-Fossilien sind 1-2 mm breit und 5 bis 13 mm lang (Bartley, 1998). Unsegmentierte Fossilien wie Protechirus, Platypholina und Vladimissa sind nicht gut verstanden. Einige der Fossilien sind segmentiert, wie z. B. Dickinsonia, das als Ringelwurm klassifiziert wird. Die kleinsten Exemplare von Dickinsonia haben einen Durchmesser von 4 mm, während die größten 1 Meter lang sind. Trotz der Länge dieser Würmer waren sie mit unter 3 mm sehr dünn. Es gibt Hinweise darauf, dass die Sauerstoffmenge in der Atmosphäre am Ende des Präkambriums nur etwa ein Zehntel der heutigen Konzentration betrug, und die extreme Dünnheit (und damit große Oberfläche) dieser Würmer könnte eine Anpassung für die Sauerstoffdiffusion gewesen sein. Diese Würmer waren häufig und weit verbreitet. Sie ernährten sich von Trümmern auf dem Meeresboden, da das Sediment oft ihren Darm füllte (Runnegar, 1982). Die einfache Natur von Dickinsonia könnte darauf hindeuten, dass es sich eher um ein Nesseltier als um einen Bilateralen handelt (Brasier, 2008).
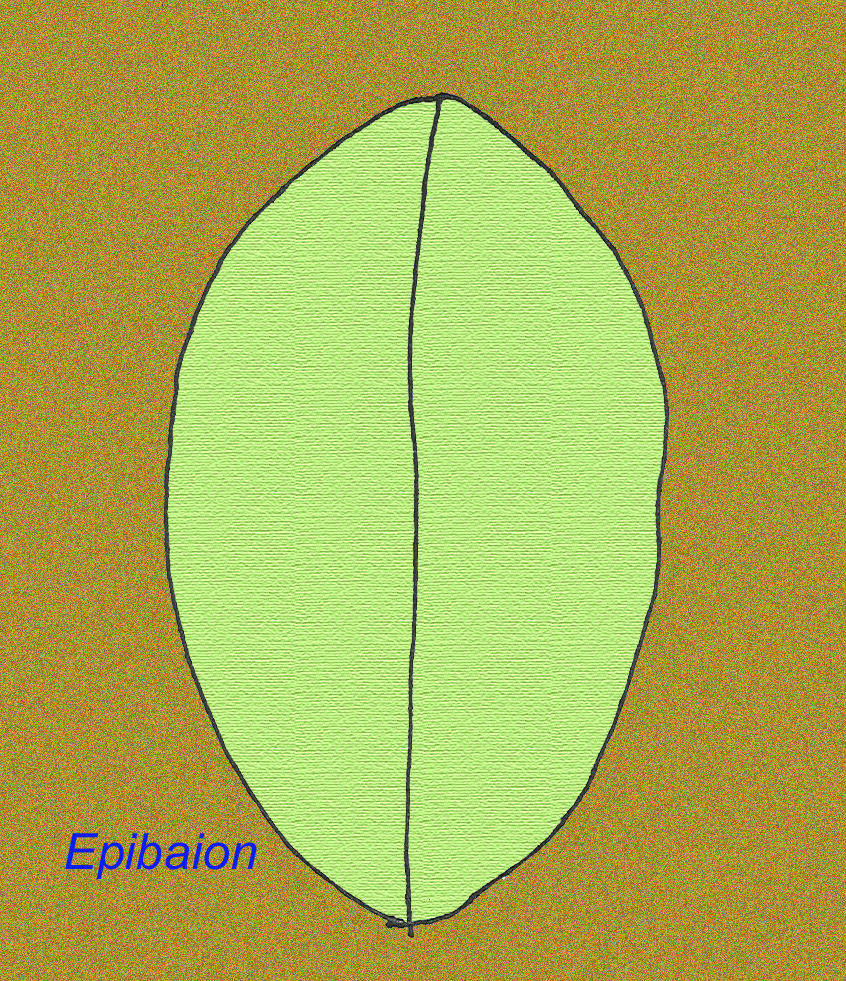
Ein großes Ediacran-Tier namens Epibaion axiferus hat Abdrücke hinterlassen, die bis zu 4,3 Meter mal 0,15 Meter messen (mit einem zweiten Exemplar, das 2,5 Meter mal 0,35 Meter misst) (Ivantsov, 2002).


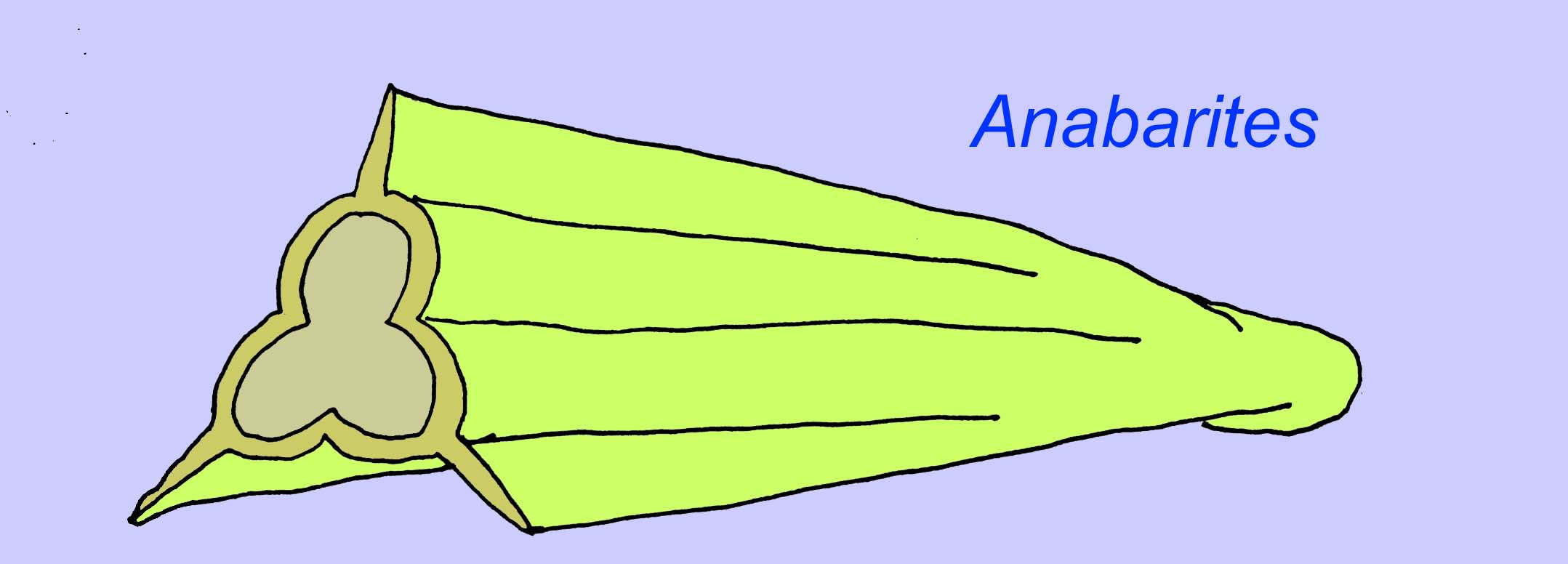
Anabaritiden sind schalenförmige Fossilien aus dem frühen Kambrium. Einige, wie z. B. Anabariten, können auf das Ende des Präkambriums datiert werden (McMenamin, 1990).
An einer Reihe von Fundstellen im Proterozoikum wurden Spurenfossilien von Wurmgruben gefunden. Diese Höhlen wurden von Würmern angelegt, die weiter entwickelt waren als Plattwürmer. (Bartley, 1998, Narbonne, 1997, Hagadorn, 1998, McMenamin, 1996). Einige moderne Nesseltiere können einfache Höhlen bilden und es ist möglich, dass einige der Höhlenspurenfossilien von Nesseltieren und nicht von Bilateralen gebildet wurden (Jensen, 2005). Einige Wurmhöhlen sind komplett mit Kot, was auf ein Verdauungssystem wie bei fortgeschritteneren Würmern schließen lässt (Morris, 1993a; Valentine 1994). Fossilien von Wurmembryonen wurden aus dem frühesten Kambrium gefunden (Bengston, 1997).
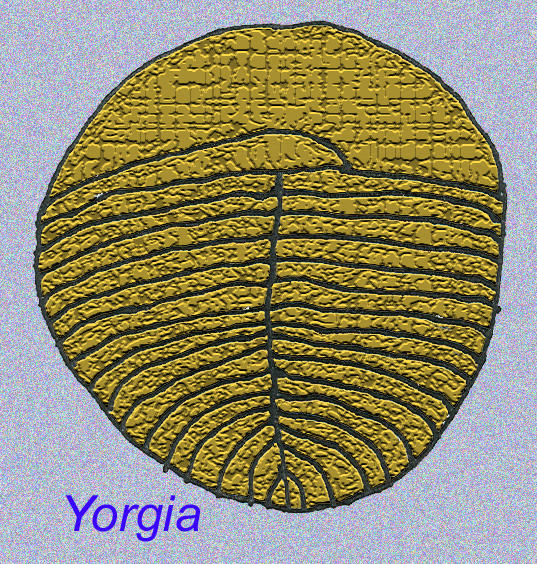
Gibt es unter den EDIACARAN-Fauna Tiere, die weiter entwickelt sind als die Würmer?
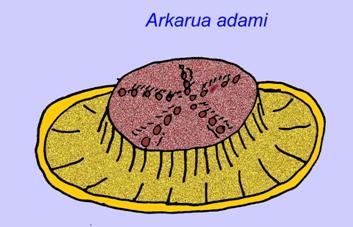
Einige genetische Analysen haben nahegelegt, dass die Trennung der Hauptgruppen der coelomaten Tiere vor dem Kambrium stattfand (Gu, 1998). Eine Reihe von Ediacaran-Fossilien wurden in coelomate Gruppen eingeordnet, was diese Schlussfolgerung unterstützt. Arkurua weist mit seiner 5-fach verzweigten Symmetrie Ähnlichkeiten zu den Stachelhäutern auf und ähnelt den primitiven Stachelhäutern des Kambriums (den Edrioasteroiden). Möglicherweise sind die 5 Strukturen Fressarme, die zu einer zentralen Rinne führen (Gehling, 1987). Zwei Gattungen (Redkinia und Spriggina) stehen möglicherweise nahe an der Basis der Gliederfüßerlinien. Tribrachidium, das auf einer seiner Oberflächen drei Anhängsel besaß, wurde in Australien und Europa gefunden (Wright, 1997b; McCall, 2006). Einige der Ediacaran-Fauna hatten einen Körperplan, der triradial (Tribrachidium, Ventagyrus), tetraradial (Conomedusies) und octaradial (Eoandromeda) war. Einige haben diese als ausgestorbene Linien von Nesseltieren interpretiert (Xiao, 2009).
Das Ediacaran-Fossil Ausia wurde als ein Urochordat klassifiziert (Xiao, 2009).
Zwei Arten, Phylozoon und Kimberella (siehe unten), könnten Verwandte von segmentierten Würmern (Anneliden) oder Mollusken sein. Kimberella hat bilaterale Symmetrie und einen möglichen Darm (Fedonkin, 1997). Kimberella scheint eine Molluske gewesen zu sein, die als schnell schwimmender Meeresräuber lebte. Die Löcher, die in die Schalen von Cloudina gebohrt wurden, wurden den Edicaran-Molluskenräubern zugeschrieben (McCall, 2006). Fossile Embryonen aus dem späten Präkambrium bildeten Polkörper, die vielen Mollusken ähneln (Chen, 2006).
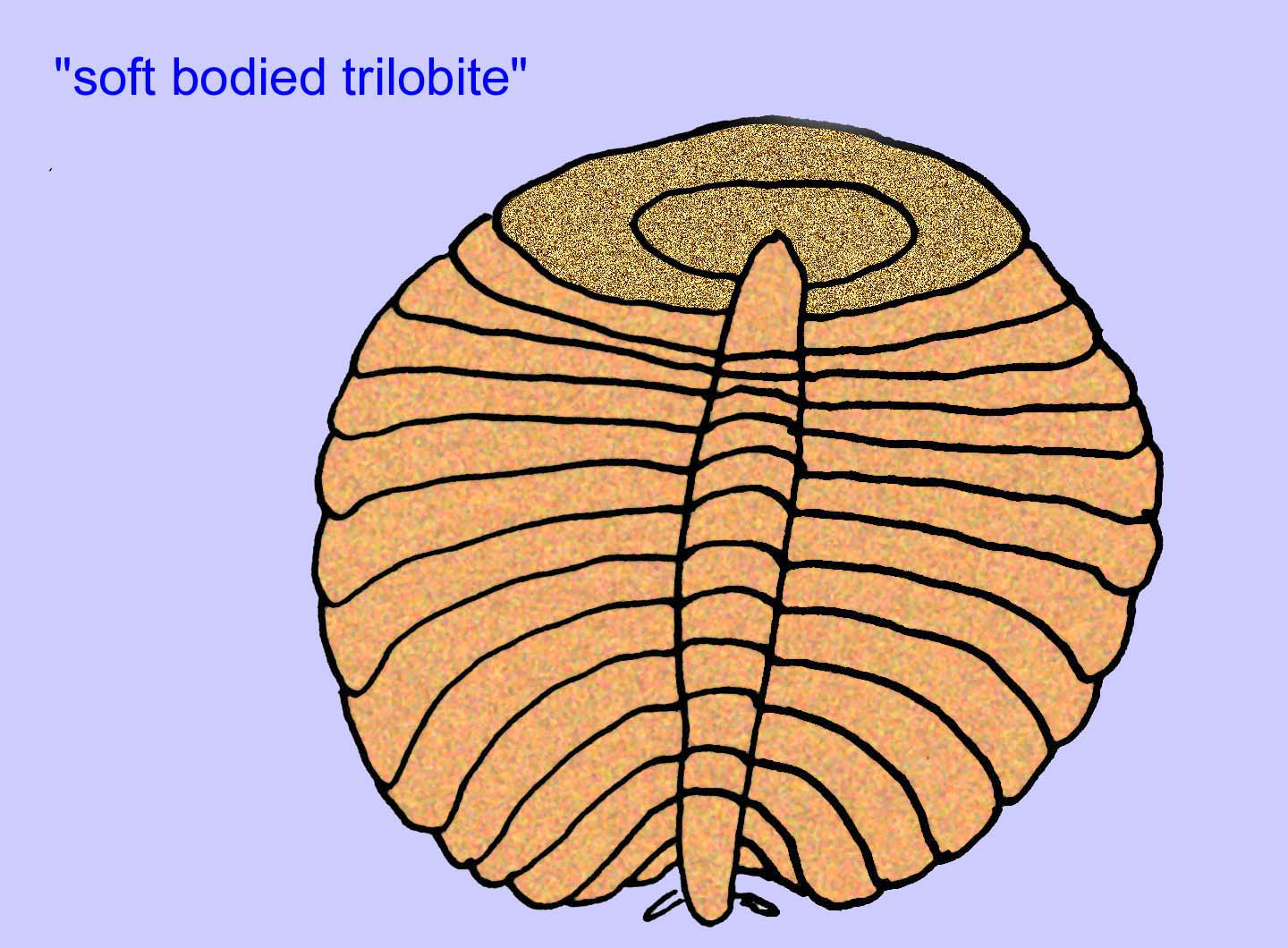
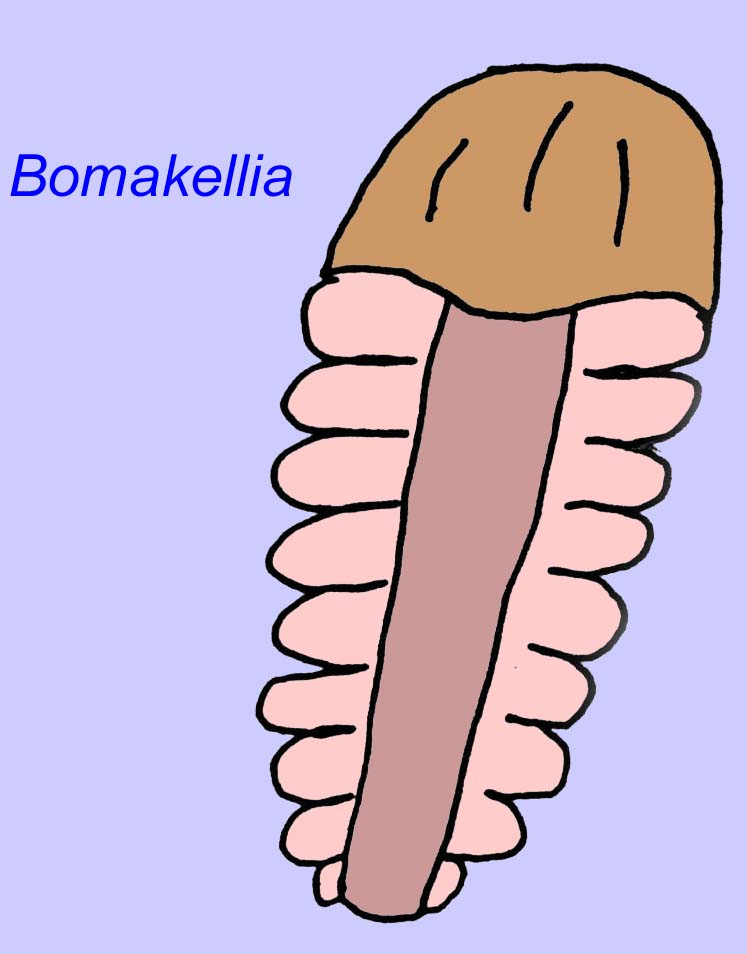
Andere Fossilien könnten mit Gliederfüßern verwandt sein, von denen einige (wie Bomakellia) eine Struktur aufweisen, die mit dem Kopfschild von Trilobiten (einer häufigen Gliederfüßerart des Paläozoikums; McCall, 2006) verglichen wurde.
Einige der Ediacaran-Fauna hatten einen Körperplan, der triradial (Tribrachidium, Ventagyrus), tetraradial (Conomedusies) und octaradial (Eoandromeda) war. Einige haben diese als ausgestorbene Linien von Nesseltieren interpretiert (Xiao, 2009).