Objectifs d’apprentissage
À la fin de cette section, vous serez en mesure de :
- Décrire la structure de l’ADN
- Décrire comment l’ADN eucaryote et procaryote est disposé dans la cellule
Dans les années 1950, Francis Crick et James Watson ont travaillé ensemble à l’Université de Cambridge, en Angleterre, pour déterminer la structure de l’ADN. D’autres scientifiques, comme Linus Pauling et Maurice Wilkins, exploraient aussi activement ce domaine. Pauling avait découvert la structure secondaire des protéines à l’aide de la cristallographie aux rayons X. La cristallographie aux rayons X est une technique qui permet de déterminer la structure de l’ADN. La cristallographie aux rayons X est une méthode qui permet d’étudier la structure moléculaire en observant les motifs formés par les rayons X projetés à travers un cristal de la substance. Ces motifs donnent des informations importantes sur la structure de la molécule en question. Dans le laboratoire de Wilkins, la chercheuse Rosalind Franklin utilisait la cristallographie aux rayons X pour comprendre la structure de l’ADN. Watson et Crick ont pu reconstituer le puzzle de la molécule d’ADN à l’aide des données de Franklin (figure 9.2). Watson et Crick disposaient également d’éléments d’information clés provenant d’autres chercheurs, comme les règles de Chargaff. Chargaff avait montré que sur les quatre types de monomères (nucléotides) présents dans une molécule d’ADN, deux types étaient toujours présents en quantités égales et que les deux autres types étaient également toujours présents en quantités égales. Cela signifie qu’ils étaient toujours appariés d’une manière ou d’une autre. En 1962, James Watson, Francis Crick et Maurice Wilkins ont reçu le prix Nobel de médecine pour leurs travaux visant à déterminer la structure de l’ADN.

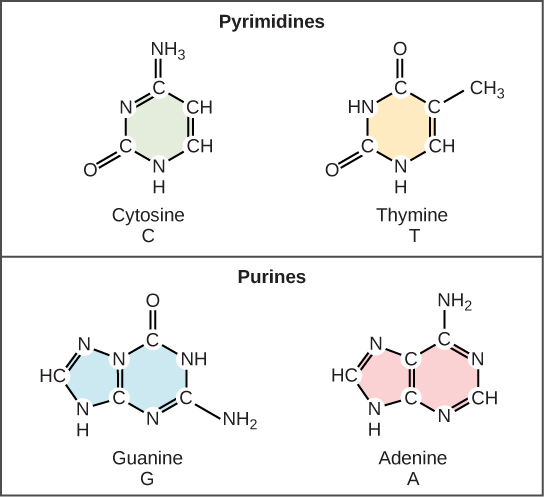
Envisageons maintenant la structure des deux types d’acides nucléiques, l’acide désoxyribonucléique (ADN) et l’acide ribonucléique (ARN). Les éléments constitutifs de l’ADN sont les nucléotides, qui sont constitués de trois parties : un désoxyribose (sucre à 5 atomes de carbone), un groupe phosphate et une base azotée (figure 9.3). Il existe quatre types de bases azotées dans l’ADN. L’adénine (A) et la guanine (G) sont des purines à deux anneaux, tandis que la cytosine (C) et la thymine (T) sont des pyrimidines plus petites à un seul anneau. Le nucléotide est nommé en fonction de la base azotée qu’il contient.
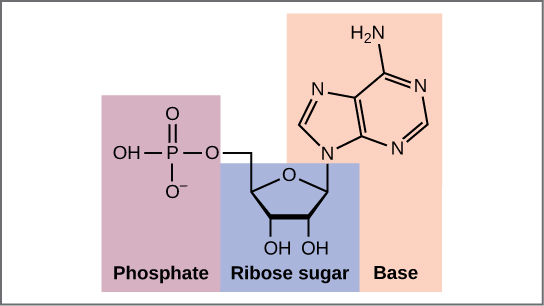
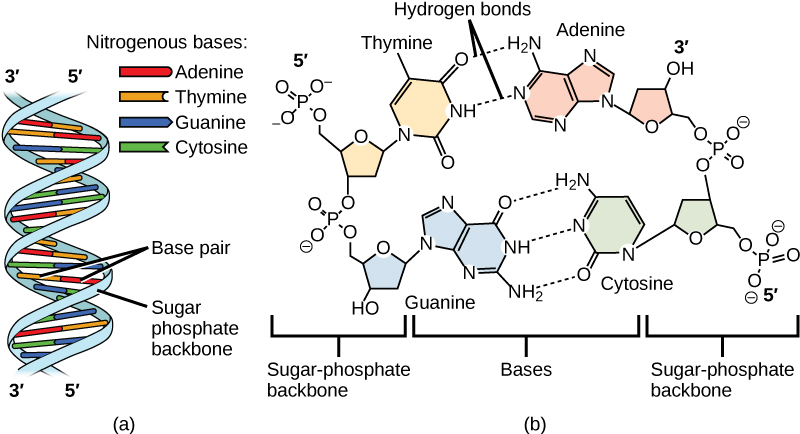
Le groupe phosphate d’un nucléotide se lie de manière covalente à la molécule de sucre du nucléotide suivant, et ainsi de suite, formant un long polymère de monomères nucléotidiques. Les groupes sucre-phosphate s’alignent en une « colonne vertébrale » pour chaque brin d’ADN, et les bases nucléotidiques partent de cette colonne vertébrale. Les atomes de carbone du sucre à cinq carbones sont numérotés dans le sens des aiguilles d’une montre à partir de l’oxygène comme 1′, 2′, 3′, 4′ et 5′ (1′ se lit comme » un premier « ). Le groupe phosphate est attaché au carbone 5′ d’un nucléotide et au carbone 3′ du nucléotide suivant. À l’état naturel, chaque molécule d’ADN est en fait composée de deux brins simples maintenus ensemble sur leur longueur par des liaisons hydrogène entre les bases.
Watson et Crick ont proposé que l’ADN soit constitué de deux brins qui s’enroulent l’un autour de l’autre pour former une hélice droite, appelée double hélice. L’appariement des bases se fait entre une purine et une pyrimidine : A s’apparie avec T, et G avec C. En d’autres termes, l’adénine et la thymine sont des paires de bases complémentaires, et la cytosine et la guanine sont également des paires de bases complémentaires. C’est la base de la règle de Chargaff ; en raison de leur complémentarité, il y a autant d’adénine que de thymine dans une molécule d’ADN et autant de guanine que de cytosine. L’adénine et la thymine sont reliées par deux liaisons hydrogène, tandis que la cytosine et la guanine sont reliées par trois liaisons hydrogène. Les deux brins sont de nature antiparallèle, c’est-à-dire qu’un brin aura le carbone 3′ du sucre en position » haute « , tandis que l’autre brin aura le carbone 5′ en position haute. Le diamètre de la double hélice de l’ADN est uniforme sur toute sa longueur car une purine (deux anneaux) s’apparie toujours avec une pyrimidine (un anneau) et leurs longueurs combinées sont toujours égales. (Figure 9.4).
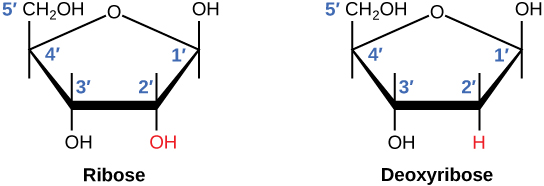
Il existe un deuxième acide nucléique dans toutes les cellules, appelé acide ribonucléique, ou ARN. Comme l’ADN, l’ARN est un polymère de nucléotides. Chacun des nucléotides de l’ARN est constitué d’une base azotée, d’un sucre à cinq atomes de carbone et d’un groupe phosphate. Dans le cas de l’ARN, le sucre à cinq carbones est le ribose, et non le désoxyribose. Le ribose possède un groupe hydroxyle sur le carbone 2′, contrairement au désoxyribose, qui ne possède qu’un atome d’hydrogène (figure 9.5).
Les nucléotides de l’ARN contiennent les bases azotées adénine, cytosine et guanine. Cependant, ils ne contiennent pas de thymine, qui est remplacée par l’uracile, symbolisé par un « U ». L’ARN existe sous la forme d’une molécule simple brin plutôt que d’une hélice double brin. Les biologistes moléculaires ont nommé plusieurs types d’ARN sur la base de leur fonction. Il s’agit notamment de l’ARN messager (ARNm), de l’ARN de transfert (ARNt) et de l’ARN ribosomal (ARNr) – des molécules qui participent à la production de protéines à partir du code de l’ADN.
Comment l’ADN est disposé dans la cellule
L’ADN est une molécule de travail ; il doit être répliqué lorsqu’une cellule est prête à se diviser, et il doit être » lu » pour produire les molécules, telles que les protéines, permettant d’assurer les fonctions de la cellule. Pour cette raison, l’ADN est protégé et emballé de manière très spécifique. En outre, les molécules d’ADN peuvent être très longues. Étirées bout à bout, les molécules d’ADN d’une seule cellule humaine atteindraient une longueur d’environ 2 mètres. Ainsi, l’ADN d’une cellule doit être emballé de manière très ordonnée pour s’adapter et fonctionner dans une structure (la cellule) qui n’est pas visible à l’œil nu. Les chromosomes des procaryotes sont beaucoup plus simples que ceux des eucaryotes par nombre de leurs caractéristiques (figure 9.6). La plupart des procaryotes contiennent un seul chromosome circulaire qui se trouve dans une zone du cytoplasme appelée nucléoïde.
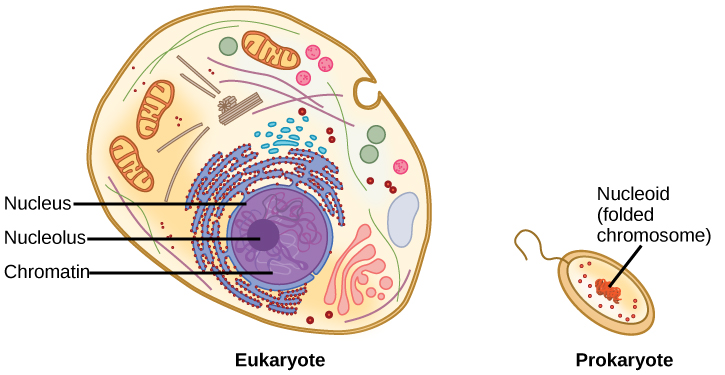
La taille du génome de l’un des procaryotes les plus étudiés, Escherichia coli, est de 4,6 millions de paires de bases, qui s’étendraient sur une distance d’environ 1,6 mm si on les étirait. Alors comment cela peut-il tenir dans une petite cellule bactérienne ? L’ADN est torsadé au-delà de la double hélice dans ce qu’on appelle le superenroulement. Certaines protéines sont connues pour être impliquées dans le superenroulement ; d’autres protéines et enzymes aident à maintenir la structure superenroulée.
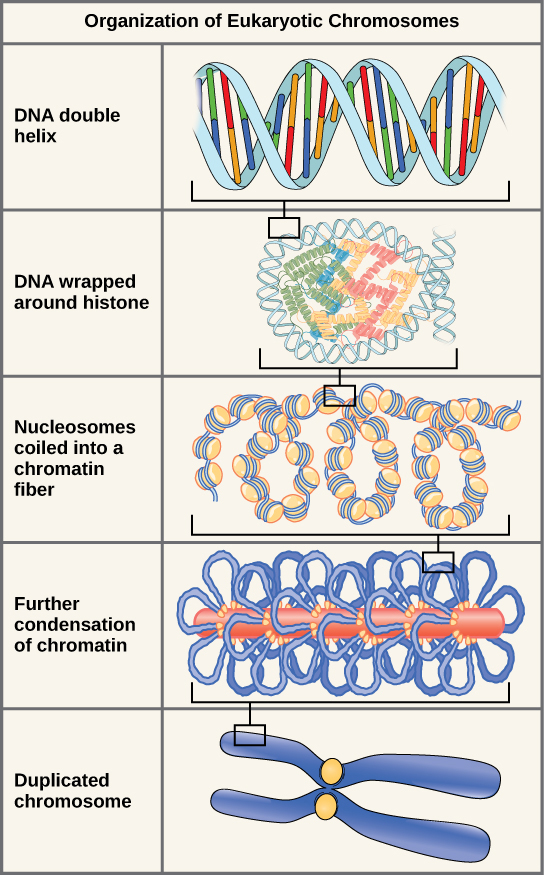
Les eucaryotes, dont les chromosomes sont chacun constitués d’une molécule d’ADN linéaire, emploient un type différent de stratégie d’emballage pour faire tenir leur ADN à l’intérieur du noyau. Au niveau le plus élémentaire, l’ADN est enroulé autour de protéines appelées histones pour former des structures appelées nucléosomes. L’ADN est étroitement enroulé autour du noyau de l’histone. Ce nucléosome est relié au suivant par un court brin d’ADN dépourvu d’histones. Cette structure est également connue sous le nom de « perles sur un fil » ; les nucléosomes sont les « perles » et les courtes longueurs d’ADN entre eux sont le « fil ». Les nucléosomes, avec leur ADN enroulé autour d’eux, s’empilent de manière compacte les uns sur les autres pour former une fibre de 30 nm de large. Cette fibre est ensuite enroulée pour former une structure plus épaisse et plus compacte. Au stade de la métaphase de la mitose, lorsque les chromosomes sont alignés au centre de la cellule, les chromosomes sont les plus compacts. Ils mesurent environ 700 nm de largeur et se trouvent en association avec des protéines d’échafaudage.
En interphase, la phase du cycle cellulaire entre les mitoses au cours de laquelle les chromosomes sont décondensés, les chromosomes eucaryotes présentent deux régions distinctes qui peuvent être distinguées par coloration. Il existe une région très dense qui présente une coloration foncée, et une région moins dense. Les régions à coloration foncée contiennent généralement des gènes qui ne sont pas actifs et se trouvent dans les régions du centromère et des télomères. Les régions à coloration claire contiennent généralement des gènes qui sont actifs, avec de l’ADN emballé autour des nucléosomes mais pas plus compacté.
Concept en action

Visionnez cette animation de l’emballage de l’ADN.
Résumé de la section
Le modèle de la structure en double hélice de l’ADN a été proposé par Watson et Crick. La molécule d’ADN est un polymère de nucléotides. Chaque nucléotide est composé d’une base azotée, d’un sucre à cinq carbones (désoxyribose) et d’un groupe phosphate. L’ADN comporte quatre bases azotées, deux purines (adénine et guanine) et deux pyrimidines (cytosine et thymine). Une molécule d’ADN est composée de deux brins. Chaque brin est composé de nucléotides liés entre eux de manière covalente entre le groupe phosphate de l’un et le sucre désoxyribose du suivant. De ce squelette partent les bases. Les bases d’un brin se lient aux bases du second brin par des liaisons hydrogène. L’adénine se lie toujours à la thymine et la cytosine se lie toujours à la guanine. Ces liaisons font que les deux brins s’enroulent l’un autour de l’autre en une forme appelée double hélice. L’acide ribonucléique (ARN) est un deuxième acide nucléique présent dans les cellules. L’ARN est un polymère monocaténaire de nucléotides. Il diffère également de l’ADN en ce qu’il contient le sucre ribose, plutôt que désoxyribose, et le nucléotide uracile plutôt que thymine. Diverses molécules d’ARN fonctionnent dans le processus de formation des protéines à partir du code génétique de l’ADN.
Les procaryotes contiennent un seul chromosome circulaire à double brin. Les eucaryotes contiennent des molécules d’ADN linéaires à double brin emballées dans des chromosomes. L’hélice d’ADN est enroulée autour de protéines pour former les nucléosomes. Les bobines de protéines sont encore plus enroulées, et au cours de la mitose et de la méiose, les chromosomes deviennent encore plus fortement enroulés pour faciliter leur déplacement. Les chromosomes ont deux régions distinctes qui peuvent être distinguées par coloration, reflétant différents degrés d’emballage et déterminées par le fait que l’ADN dans une région est exprimé (euchromatine) ou non (hétérochromatine).
Glossaire
Désoxyribose : une molécule de sucre à cinq carbones avec un atome d’hydrogène plutôt qu’un groupe hydroxyle en position 2′ ; le composant sucre des nucléotides de l’ADN
double hélice : la forme moléculaire de l’ADN dans laquelle deux brins de nucléotides s’enroulent l’un autour de l’autre en forme de spirale
base azotée : une molécule contenant de l’azote qui agit comme une base ; se réfère souvent à l’un des composants purine ou pyrimidine des acides nucléiques
groupe phosphate : un groupe moléculaire constitué d’un atome de phosphore central lié à quatre atomes d’oxygène
.