Les premiers animaux
Les êtres vivants les plus primitifs sont unicellulaires : l’organisme entier est composé d’une seule cellule. Certains organismes peuvent vivre en tant que cellules individuelles ou en tant que groupes de cellules dans une colonie. Si les cellules d’une colonie sont séparées, elles peuvent continuer à vivre en tant que cellules individuelles ou former une nouvelle colonie. Aucune cellule n’est spécialisée dans une tâche spécifique. L’algue moderne Volvox en est un bon exemple : la photo ci-contre représente une colonie de Volvox.
La multicellularité a évolué indépendamment quelques dizaines de foisdans les plantes, les algues rouges, les algues brunes, les champignons et les animaux. Par exemple, les algues rouges du groupe des Volvocales comprennent des espèces unicellulaires (Chlamydomonas), des espèces quadricellulaires généralisées (Basichlamys) et des espèces qui forment des colonies pouvant comprendre des colonies de 50 000 cellules (Volvox) (Sachs, 2008).
Si beaucoup d’algues vivent dans ces colonies simples, certaines algues ont en fait des cellules qui se spécialisent pour effectuer différentes tâches (elles ont des tissus qui ont une fonction de racine et une fonction de feuille).
Les premiers fossiles d’algues sont connus il y a environ 2,1 milliards d’années et, vers 1 milliard d’années, on observe une augmentation de la diversité des espèces d’algues connues. Les algues eucaryotes semblent s’être diversifiées brusquement il y a environ 600 millions d’années, suite aux glaciations de la fin du Protérozoïque (Cavalier-Smith, 2000).
Dès 570 millions d’années, les algues sont connues avec une variété de caractéristiques anatomiques et reproductives (Xiao, 1998). Le registre fossile a préservé une diversité d’algues préhistoriques telles que celles représentées ci-dessous.
FONGES
De nombreux fossiles pré-édiacariens représentent des colonies multicellulaires simples de bactéries, des algues eucaryotes (y compris des algues vertes et rouges), et des formes qui présentent des ressemblances avec des champignons. (Butterfield, 2009 ; Xiao, 2009).
Des champignons carnivores formant des anneaux hyphaux pour s’attaquer aux nématodes ont été observés dans de l’ambre vieux de 100 millions d’années (Schmidt, 2007).
Les premiers animaux
La faune édiacarienne est un assemblage d’organismes nommé d’après les collines d’Ediacara en Australie où ils ont été découverts pour la première fois. Des fossiles édiacariens sont connus dans plus de 30 pays et sur tous les continents, à l’exception de l’Antarctique et de l’Amérique du Sud (De, 2005 ; McCall, 2006).Plusieurs milliers de spécimens, comprenant plus de 270 espèces, ont été décrits dans la faune édiacarienne et beaucoup restent à décrire (Fedonkin, 1990 ; Minnerop, 2000 ; Waggoner, 1998 ; Shen, 2008). Le premier animal édiacarien a été décrit en 1872, suivi d’une deuxième découverte en 1908-14 (McCall, 2006).
- Ils sont anciens
Les strates fossiles qui ont été datées à plus de 1,2 milliard d’années contiennent des animaux – non seulement elles contiennent des fossiles d’organismes à corps mou radial, mais elles contiennent des terriers qui semblent avoir été faits par des vers producteurs de mucus (Rasmussen, 2002 ; McCall, 2006). Il existe un site en Inde dont les terriers de vers ont été datés de 1,1 milliard d’années, bien que certains contestent cette date. Récemment, un autre site présentant des signes apparents d’animaux primitifs (parents des méduses) et des terriers de vers a été daté à 1,2 milliard d’années au plus jeune (Seilacher, 1998 ; Kerr, 1998b ; Morris, 1993). D’autres fossiles édiacariens sont connus il y a 610 à 600 millions d’années et des assemblages d’organismes plus diversifiés sont connus il y a 555,3 millions d’années (McMenamin, 1996, Minnerop, 2000).Les strates de roches précambriennes datant de 580 millions d’années possèdent des fossiles de petits bilatériens ; des animaux bilatériens plus grands sont présents après des dates de 555 millions d’années (Condon, 2005).Ils ont été largement remplacés par les animaux de » l’explosion cambrienne » qui a commencé il y a environ 545 millions d’années. La période édiacarienne est définie à 635 millions d’années après la glaciation de la Terre boule de neige marinoise (bien que la glaciation de Gaskiers, il y a 562 millions d’années, se soit produite au cours de la période édiacarienne) (Xiao, 2009).
Les comparaisons moléculaires d’organismes modernes soutiennent également la conclusion selon laquelle les animaux avaient produit une variété de lignées avant le Cambrien. Par exemple, une étude portant sur 129 protéines suggère que les eucaryotes sont apparus entre 950 et 1 260 millions d’années, les animaux entre 761 et 957 millions d’années et les lignées de cœlomates se sont séparées entre 642 et 761 millions d’années (Douzery, 2004). D’autres analyses ont donné des résultats similaires (Peterson, 2004). D’autres estimations moléculaires de l’origine des animaux eumétazoaires (cnidaires et bilatériens) fournies par des études d’horloges moléculaires ont varié de 570 millions d’années à 1 100 millions d’années. L’origine des animaux bilatériens a été estimée à 580 millions d’années (Peterson, 2005). Des fossiles d’animaux trouvés en Australie, en Inde et au Texas ont été datés de plus d’un milliard d’années et un animal segmenté de Russie a été daté de 890 millions à un milliard d’années (De, 2005). En raison des découvertes passionnantes d’animaux précambriens, une nouvelle période a été ajoutée à l’échelle des temps géologiques, ce qui est la première fois qu’une nouvelle période a été définie stratigraphiquement depuis 1891 (lorsque le Carbonifère a été divisé). La période édiacarienne est définie comme la période comprise entre 630 millions d’années et le début du Cambrien, il y a environ 542 millions d’années (Knoll, 2004).
Quelle est la signification de ces dates ? Jusqu’à récemment, on pensait qu’une diversité d’animaux complexes était apparue soudainement lors de « l’explosion cambrienne » débutant il y a environ 545 millions d’années. Nous savons maintenant que les animaux ont existé pendant au moins 60 millions d’années avant cela et peut-être jusqu’à 600 millions d’années avant le Cambrien. Soixante millions d’années ne semblent peut-être pas si longues au vu des vastes étendues de temps du Précambrien, mais une grande partie de l’évolution animale peut se produire au cours d’une telle période. Il a fallu environ soixante millions d’années pour que les poissons rhipidistiens se transforment en premiers amphibiens et environ soixante millions d’années pour la diversification ultérieure de ces amphibiens en diverses lignées, dont les premiers reptiles. Une période de soixante millions d’années commençant à la fin du Permien comprendrait deux extinctions massives. Elle commence par un monde dominé par les reptiles synapsides et les archosaures primitifs et se termine au Jurassique avec ses diverses lignées de dinosaures, de ptérosaures et de reptiles marins. En soixante millions d’années environ, les mammifères primitifs qui ont survécu à l’extinction des dinosaures rayonnent dans les divers groupes fossiles et vivants du Cénozoïque.


2) ILS SONT DES ANIMAUX SIMPLES
La plupart des organismes édiacariens mesurent moins d’un pouce, mais certains font plusieurs pieds de long. Ils sont bizarres et la plupart sont très différents de tout ce qui vit aujourd’hui (beaucoup se réfèrent à eux comme au groupe Vendobionta). Presque tous n’ont pas de tête ou de queue, d’intérieur ou d’extérieur, d’avant ou d’arrière, de dents ou d’yeux. La plupart n’ont pas de système circulatoire, nerveux ou digestif évident, ni d’organes internes d’aucune sorte. La plupart sont très minces et plats – une condition qui serait essentielle pour les animaux sans système circulatoire, respiratoire, digestif ou excréteur. Certains sites présentent une diversité d’animaux, avec jusqu’à 30 taxons différents (Buss, 1994).
Qu’est-ce que c’est ? Il y a eu une diversité d’opinions concernant ces organismes dans le passé : certains les ont classés comme des plantes (la conclusion à partir de la Première Guerre mondiale), de très grands organismes unicellulaires, des lichens uniques, un ensemble complètement éteint d’animaux, et un groupe d’animaux qui comprend les ancêtres des animaux modernes. On pensait à l’origine qu’ils s’étaient éteints bien avant l’apparition des groupes d’animaux modernes, mais il est maintenant évident que certains groupes ont survécu jusqu’au Cambrien et que certains pourraient représenter des fossiles d’animaux non métazoaires (Droser, 2006). La densité des individus de l’animal discoïdal Aspidella indique que la biomasse des animaux de fond au Précambrien aurait pu approcher celle observée dans les communautés modernes (Gehling, 2000).
De nombreux fossiles pré-édiacariens représentent des colonies multicellulaires simples de bactéries, d’algues eucaryotes (y compris des algues vertes et rouges), et des formes qui présentent des ressemblances avec des champignons. Certains fossiles néoprotérozoïques ont été interprétés comme des spicules et des larves d’éponges (Butterfield, 2009).
Les éponges, qui sont l’un des types d’animaux les plus simples, ont été identifiées dans des sédiments édiacariens (comme le genre Palaeophragmodictya) qui sont similaires à certaines éponges paléozoïques. Ces éponges pouvaient mesurer de 1,5 à 10 cm de diamètre (Gehling, 1996 ; Kerr, 1998a). Des spicules d’éponges sont présentes dans les roches édiacariennes (Xiao, 2000).
Certains fossiles édiacariens ont été interprétés comme des champignons et des animaux basaux plus primitifs que les éponges (Xiao, 2009).
CNIDARIENS
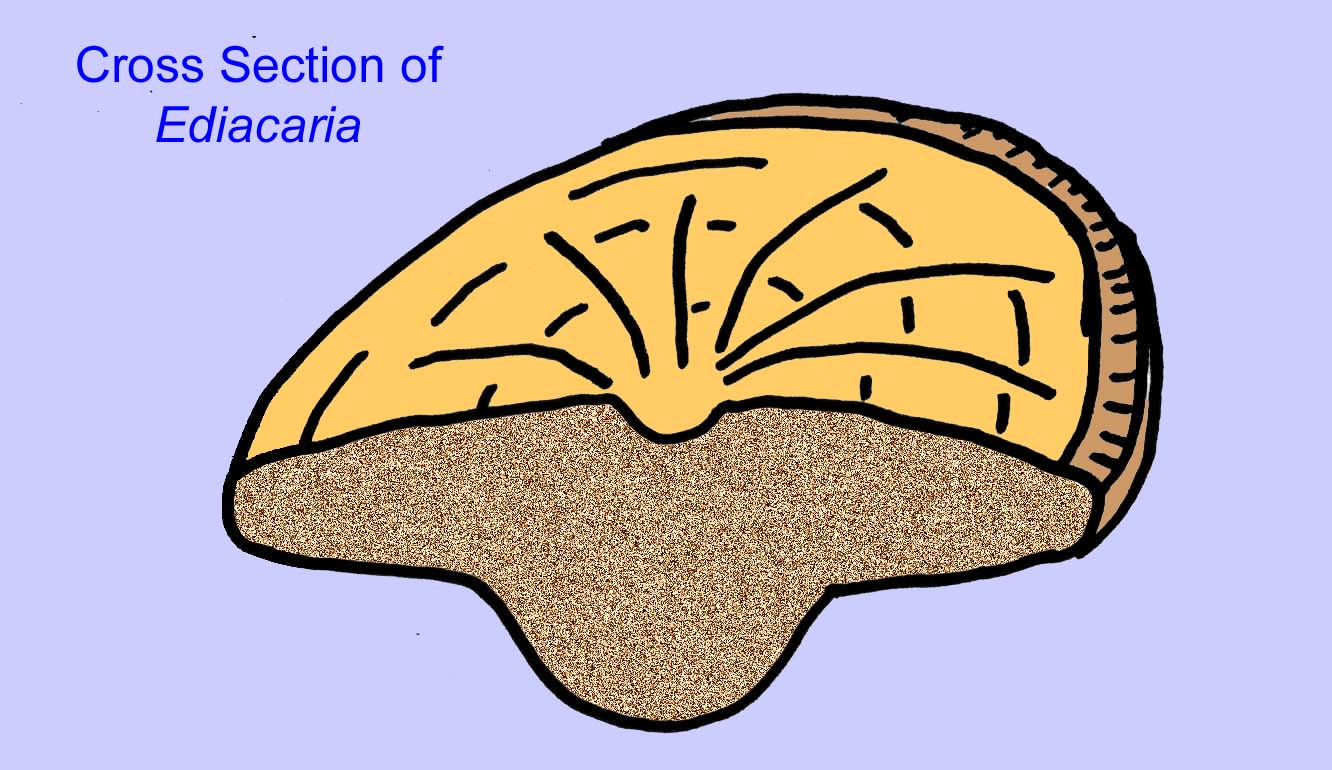
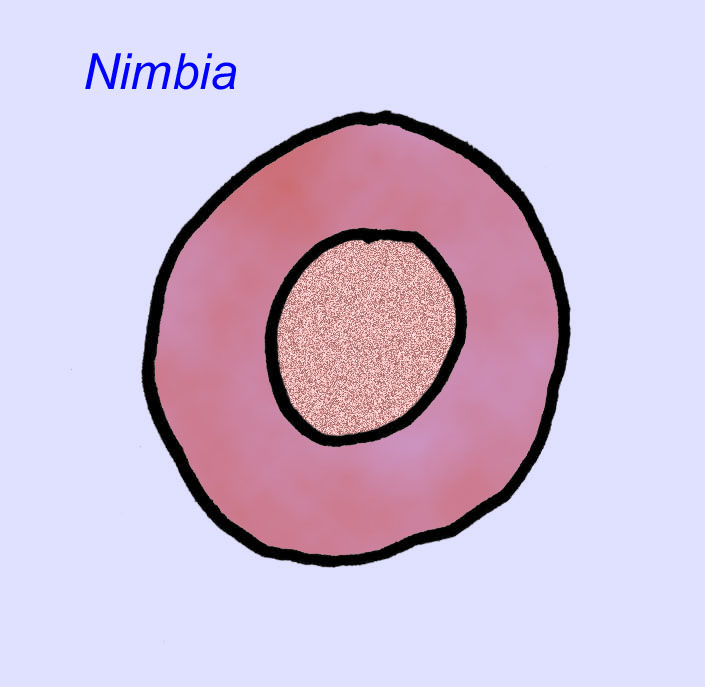
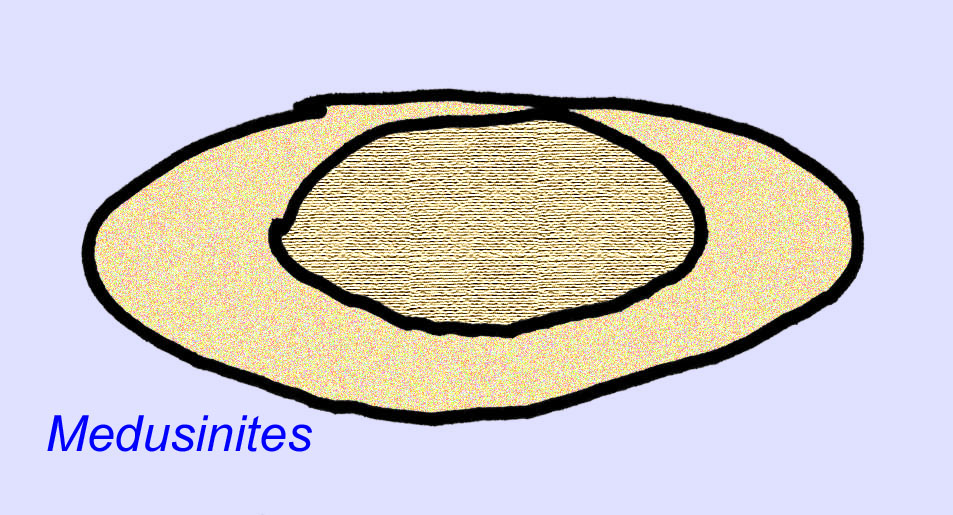
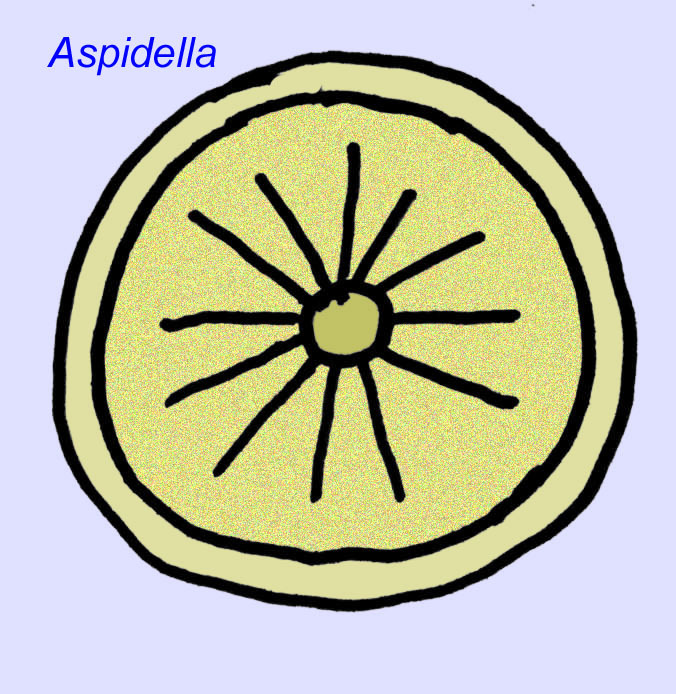
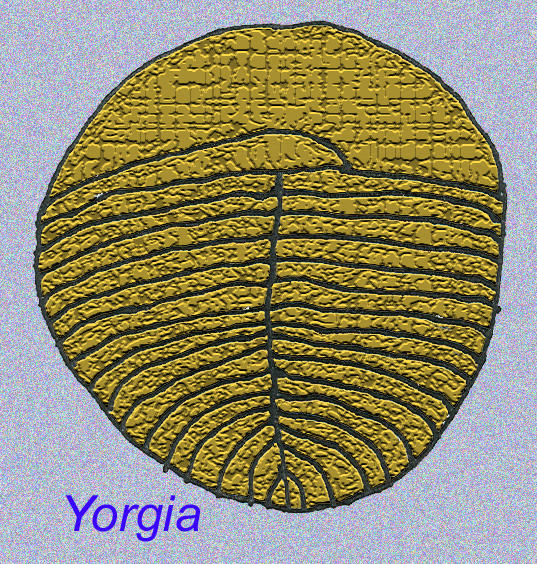
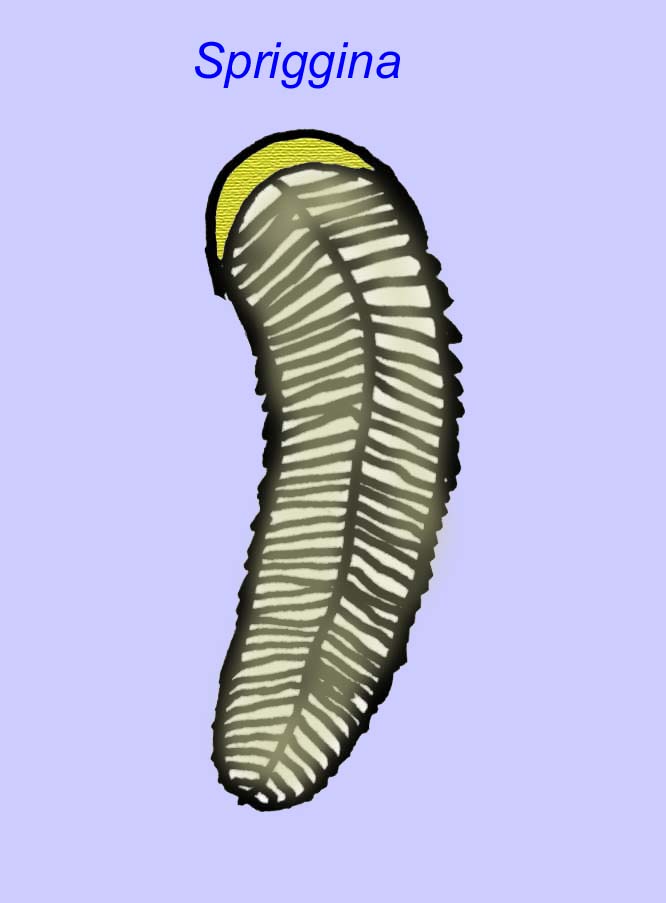
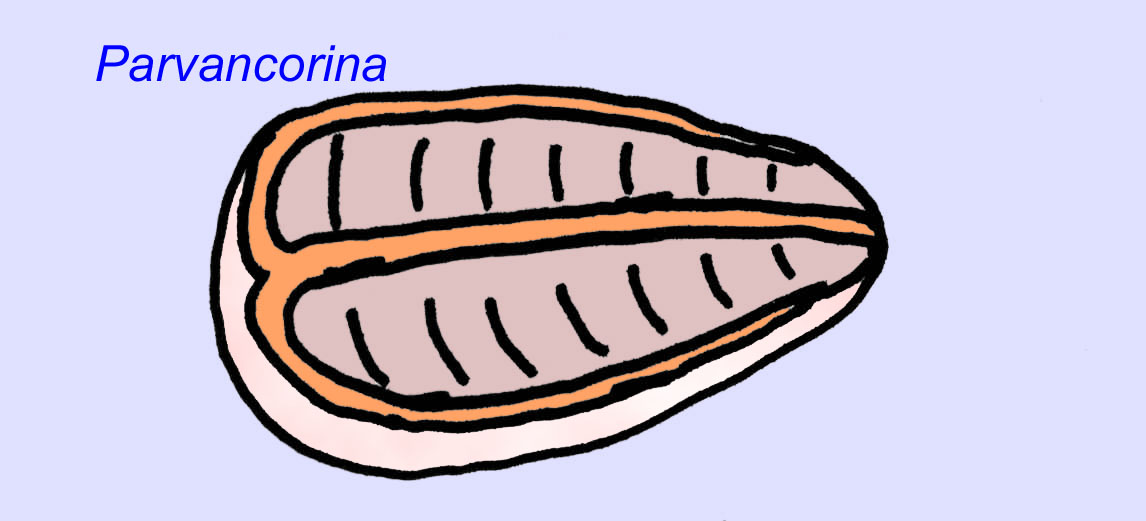
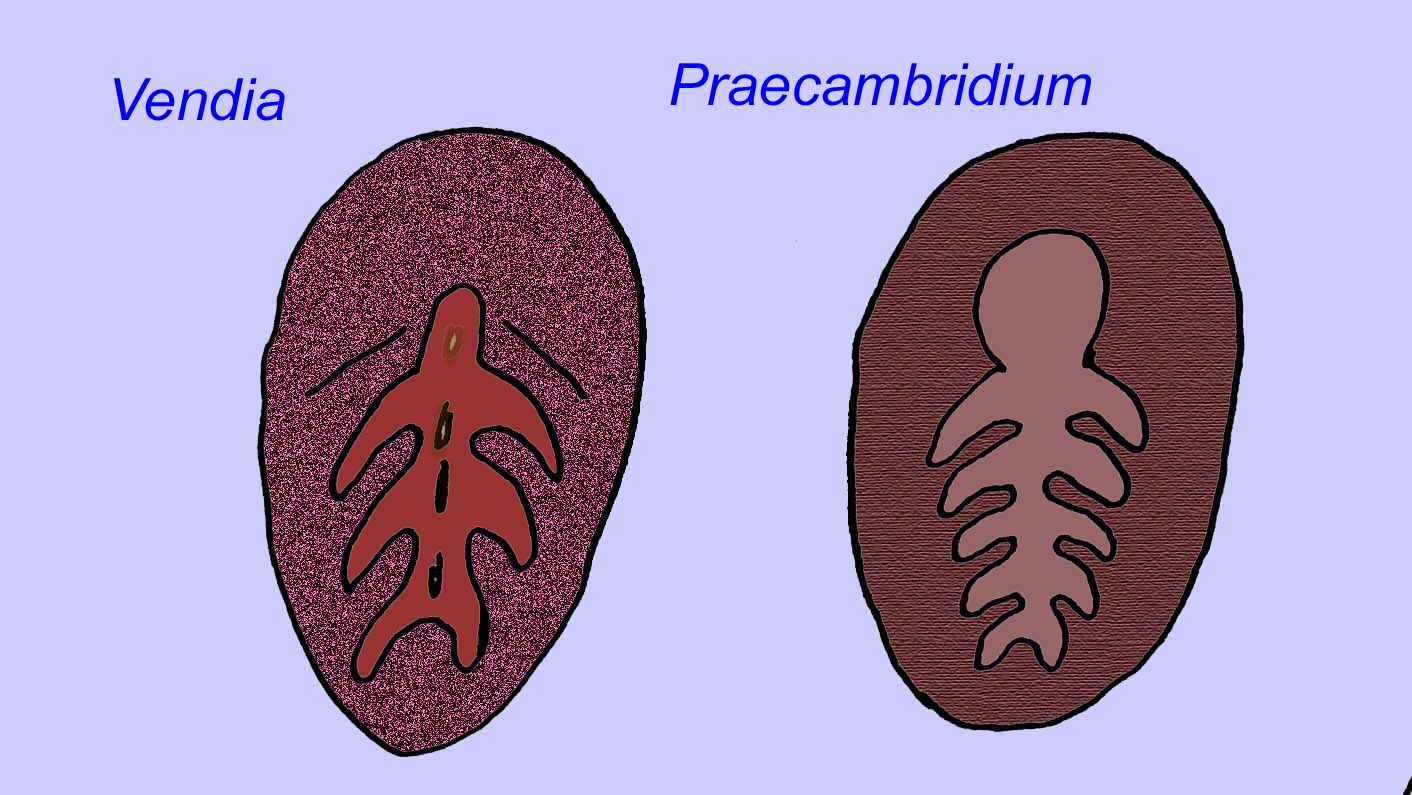
Il existe un consensus général sur le fait que de nombreux fossiles d’Édiacaran doivent être classés comme des cnidaires précoces, un groupe d’animaux qui comprend les méduses modernes, les stylos de mer et les éventails de mer. Certaines formes (comme Ediacaria et Nimbia) sont radialement symétriques et superficiellement similaires aux cnidaires, mais ne présentent aucun signe de bouche, ce qui peut être une raison pour les exclure de ce groupe. (Il n’est pas certain que les premiers cnidaires auraient eu une bouche comme toutes les formes ultérieures). D’autres, comme Albumares brunsae, possèdent un système gastrovasculaire similaire à celui des cnidaires (Lipps, 1992 ; Sokolov, 1985). Beaucoup ont une structure discoïdale centrale sur un côté de leur corps plat et rond (Crimes, 1995 ; Bertrand-Sarfati, 1995). Certains animaux d’Ediacara (Spriggina, Parvancorina, et Praecambridium) ont des distinctions entre leurs côtés supérieurs et inférieurs (postérieurs et antérieurs). Certains, comme Ediacaria, avaient des lignes radiales et un épaississement périphérique (Crimes, 1995). Certains des cnidaires de l’Édiacarien ont été classés comme une sous-classe distincte mais éteinte (McCall, 2006).
Bien que ces animaux ne possédaient pas de squelette minéralisé, la plupart ou même tous (comme Cyclomedusa et Ernietta) possédaient une sorte de squelette rigide capable de faire une empreinte dans la boue. Le type de tissu qui composait le squelette interne n’est pas connu (Crimes, 1995 ; Dzik, 1999).
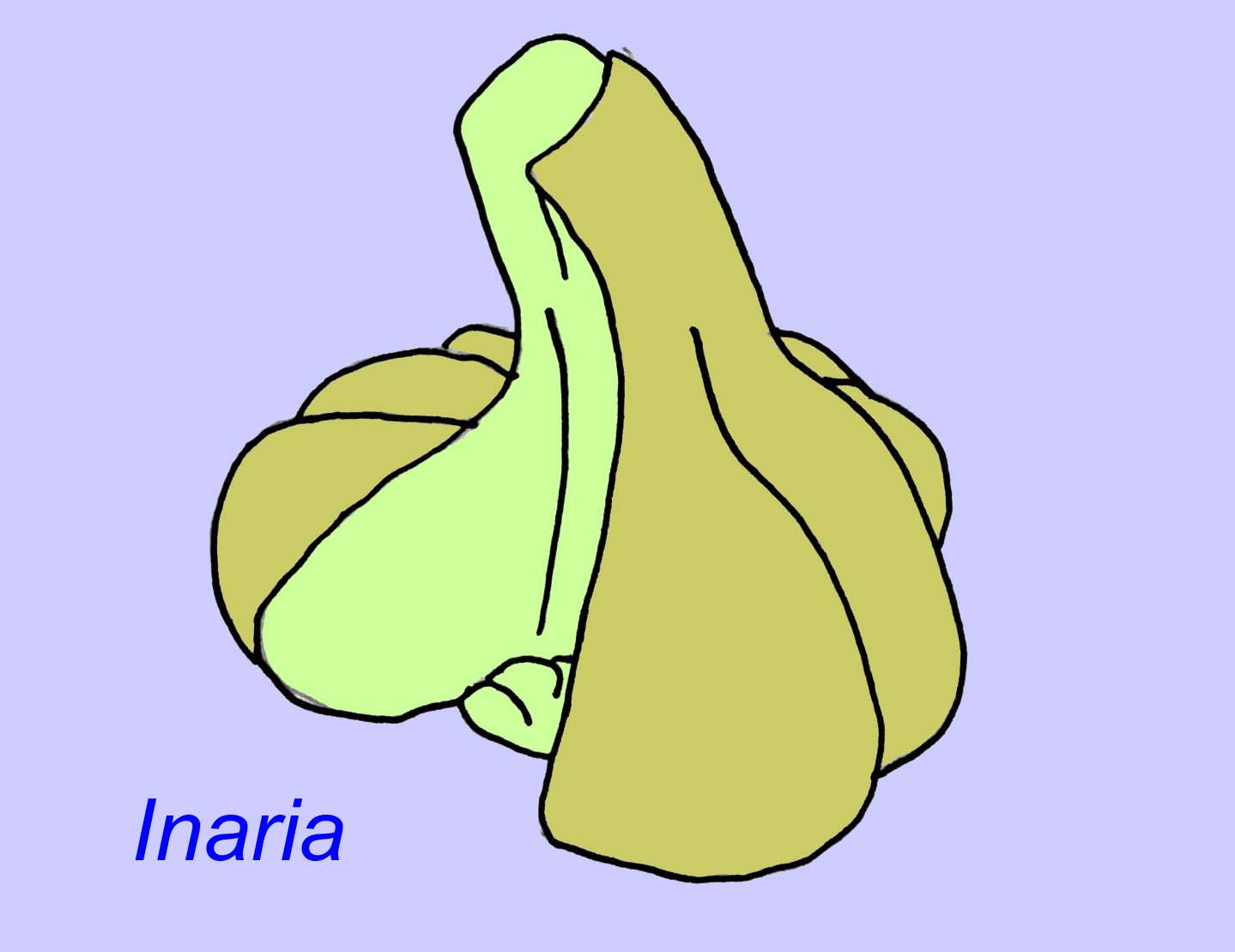
Mackenzia semble également être un cnidaire (même si ce n’est pas le cas, c’est presque certainement un animal métazoaire). Il existe des preuves indirectes de bolus alimentaires dans une cavité digestive et de musculature (Morris, 1993a). Sinocyclocyclicus pourrait être un cnidaire basal (ou même un métazoaire basal) dont le bourgeonnement apical est similaire à celui de certains cnidaires (comme les coraux tabulés) (Xiao, 2000). Inaria était un cnidaire de la classe des Anthozoaires doté d’un rebord buccal, d’un pharynx et d’une chambre qui a pu accueillir des organismes photosynthétisants comme on en trouve dans de nombreux coraux modernes (McMenaman, 1990, Gehling, 1988).
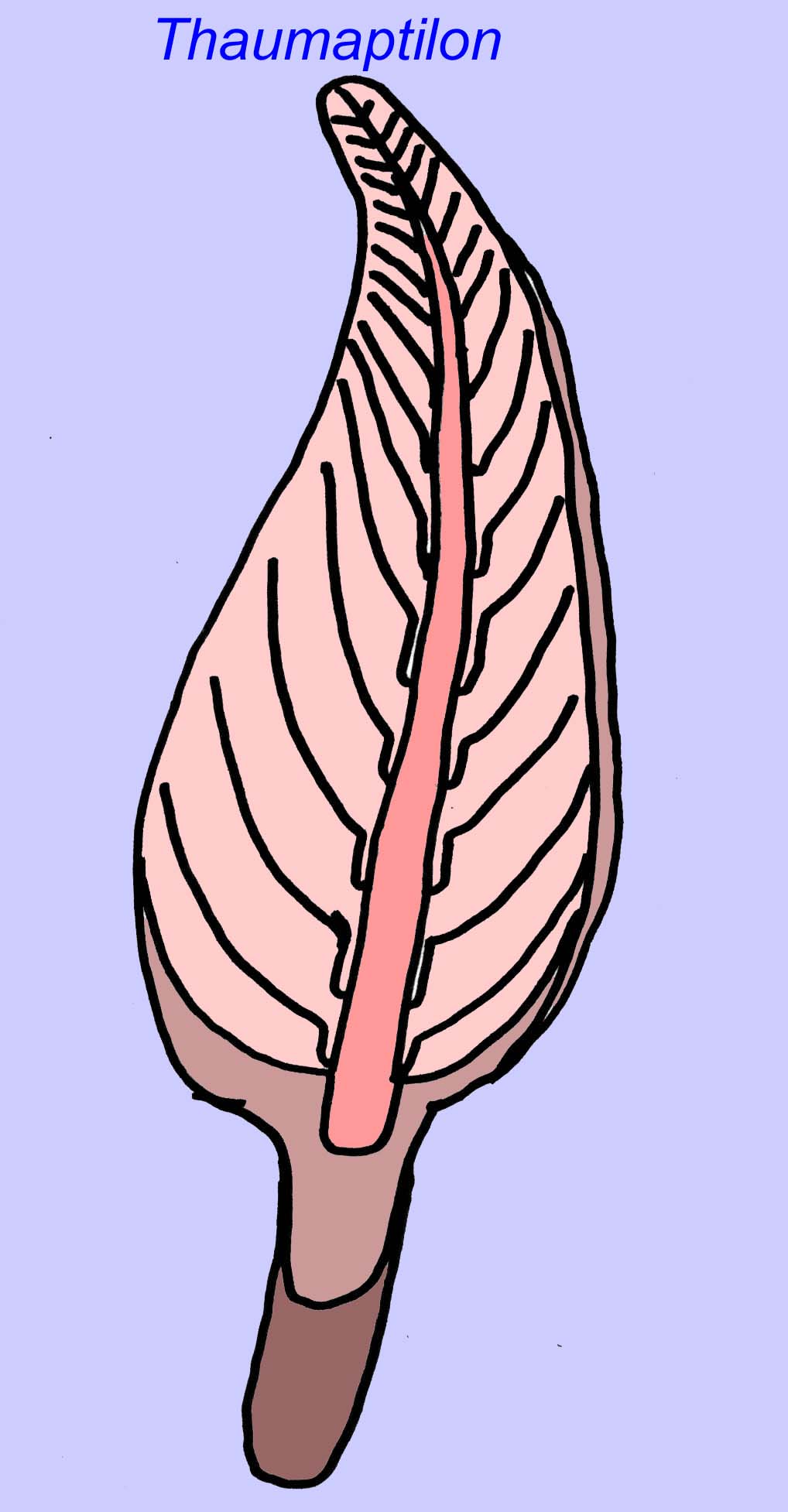
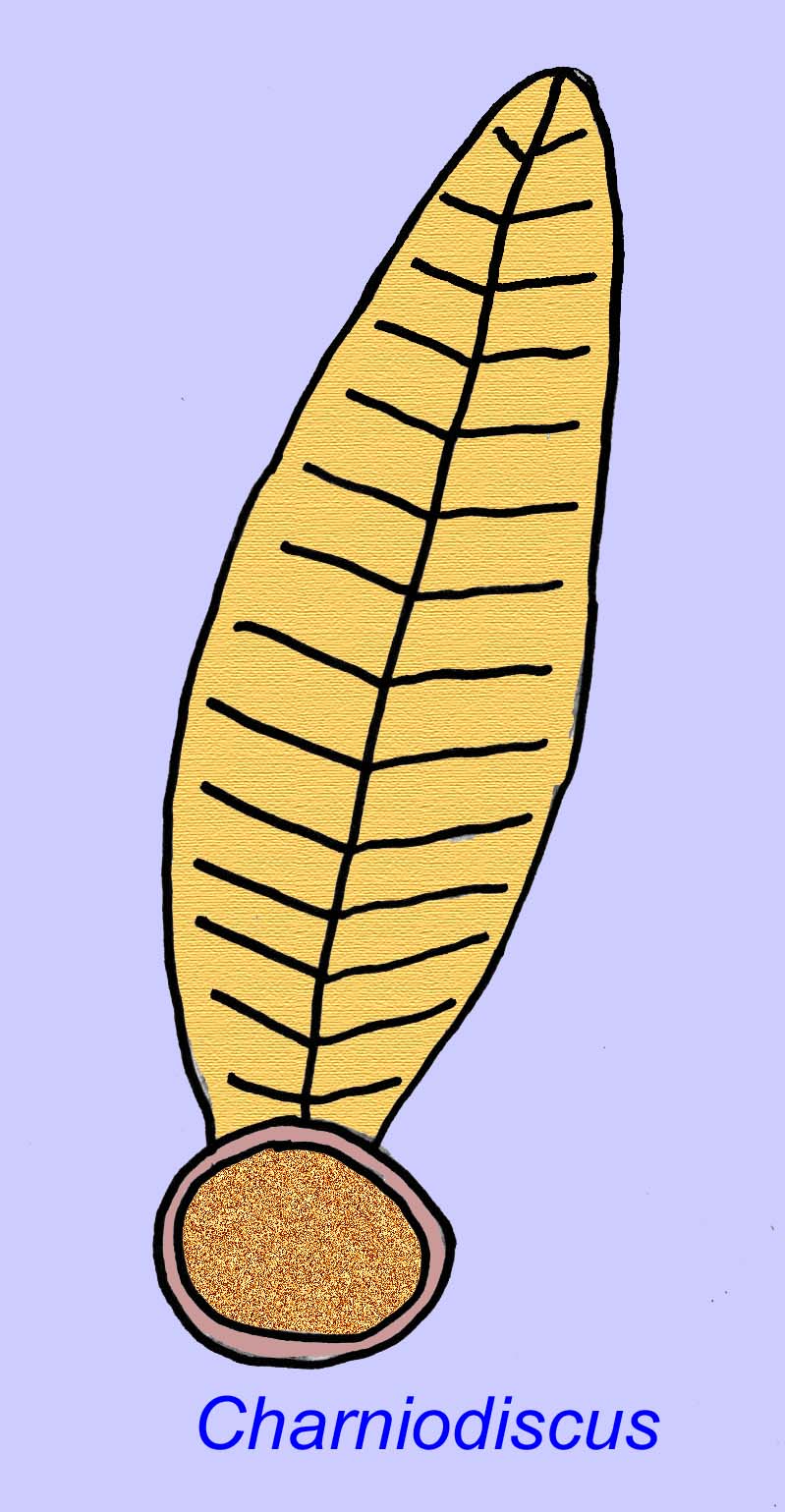
Thaumaptilon était un animal de type frondeur qui semble apparenté aux stylos de mer modernes. Il se serait nourri de débris particulaires et aurait utilisé un crampon pour se fixer au fond (Morris, 1993a ; 1993b). La fronde de Charniodiscus pouvait atteindre une longueur de 1,2 mètre (Sokolov, 1990).
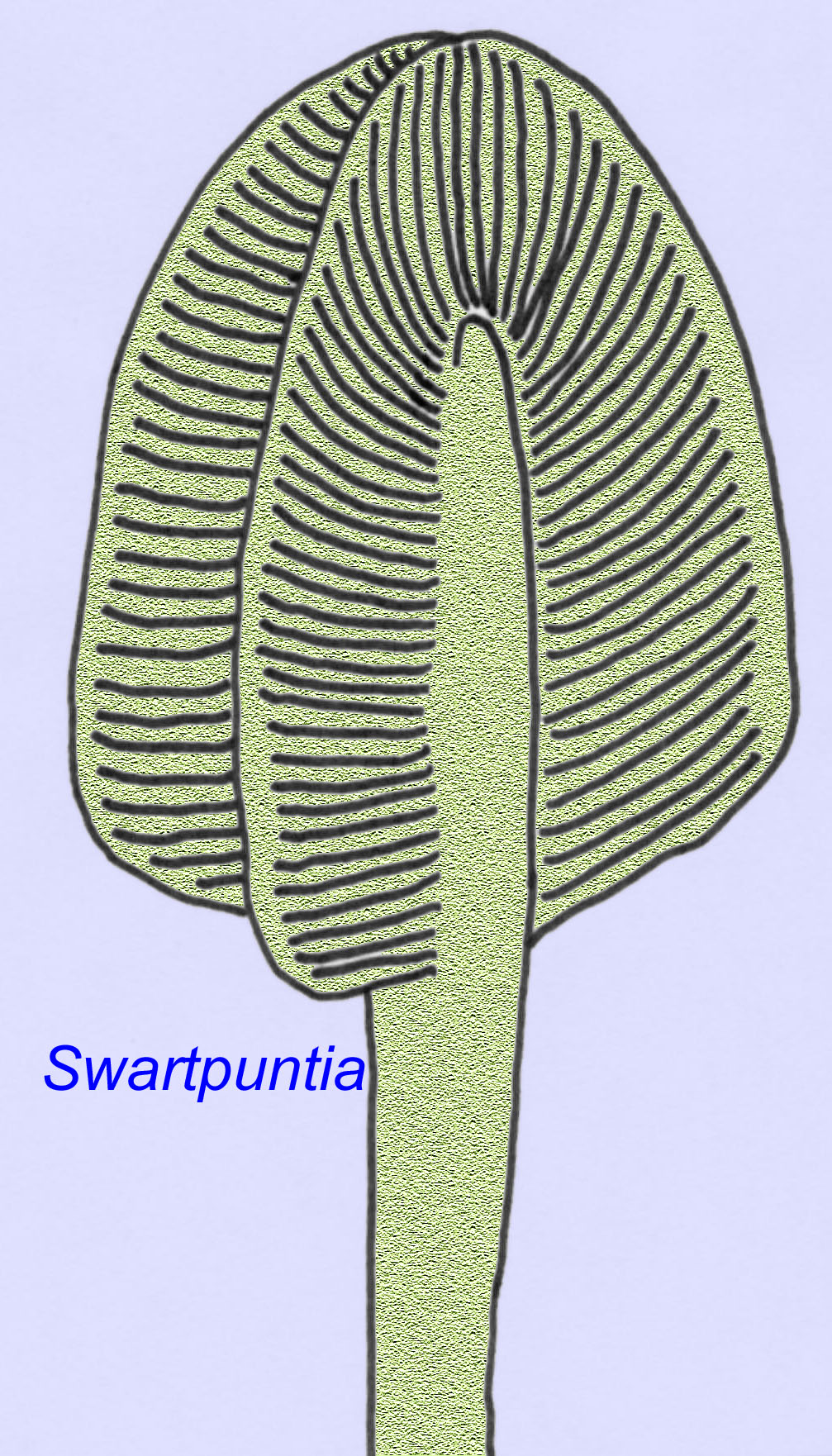
De nombreux animaux de l’Édiacarien (tels que Pteridinium, Swartpuntia, Charnia et Charniodiscus) possèdent un pédoncule vertical relié à des pétaloïdes segmentés.
Un spécimen de Charnodiscus mesurait 1,2 mètre de long.
L’anatomie microscopique de Swartpuntia est très similaire à celle de Dickinsonia, un ver précoce (Narbonne, 1997 ; McCall, 2006). Des embryons fossilisés qui semblent appartenir à des animaux bilatériens sont connus depuis 570 millions d’années (Xiao, 1998).

Les vers édiacariens sont connus en Asie, Europe, d’Afrique, d’Australie et d’Amérique du Nord (McCall, 2006). Un certain nombre d’animaux édiacariens étaient tubulaires comme Archaeichnium, Cloudina, Corumbella et Onuphionella. Alors qu’Onuphionella a été comparé à des vers annélides, Corumbella a été comparé aux tubes construits par les polypes de certains cnidaires. Alors qu’Archaeichnium était flexible, Cloudina était recouvert d’un squelette minéralisé. Certains tubes de Cloudina (longueur maximale inférieure à 4 cm) étaient annulés et d’autres étaient lisses, bien que cela puisse refléter des différences de préservation (Hagadorn, 2000). Cloudina est le premier animal connu à posséder un squelette minéralisé et 3 % des spécimens présentent des trous percés qui indiquent une prédation. Il se peut que cette prédation ait encouragé les structures plus dures typiques de tant d’animaux du Cambrien (Bengston, 1992). Cloudina est connu du Précambrien de l’Amérique du Sud, de l’Europe, du Moyen-Orient et de l’Asie du Sud-Est et des parents sont connus du Cambrien précoce (avant la propagation des trilobites) en Amérique du Nord (Grant, 1990). Les fossiles de Cloudina sont connus depuis au moins 6 millions d’années avant le début de la période cambrienne (Condon, 2005).


En plus des deux espèces reconnues de Cloudina, un certain nombre d’autres espèces fossiles possédaient des coquilles calcifiées au Précambrien supérieur (McCall, 2006). Cloudina, Namacalanthus et Conotubus sont des animaux précambriens squelettiques qui sont également connus au Cambrien précoce (Shu, 2008).
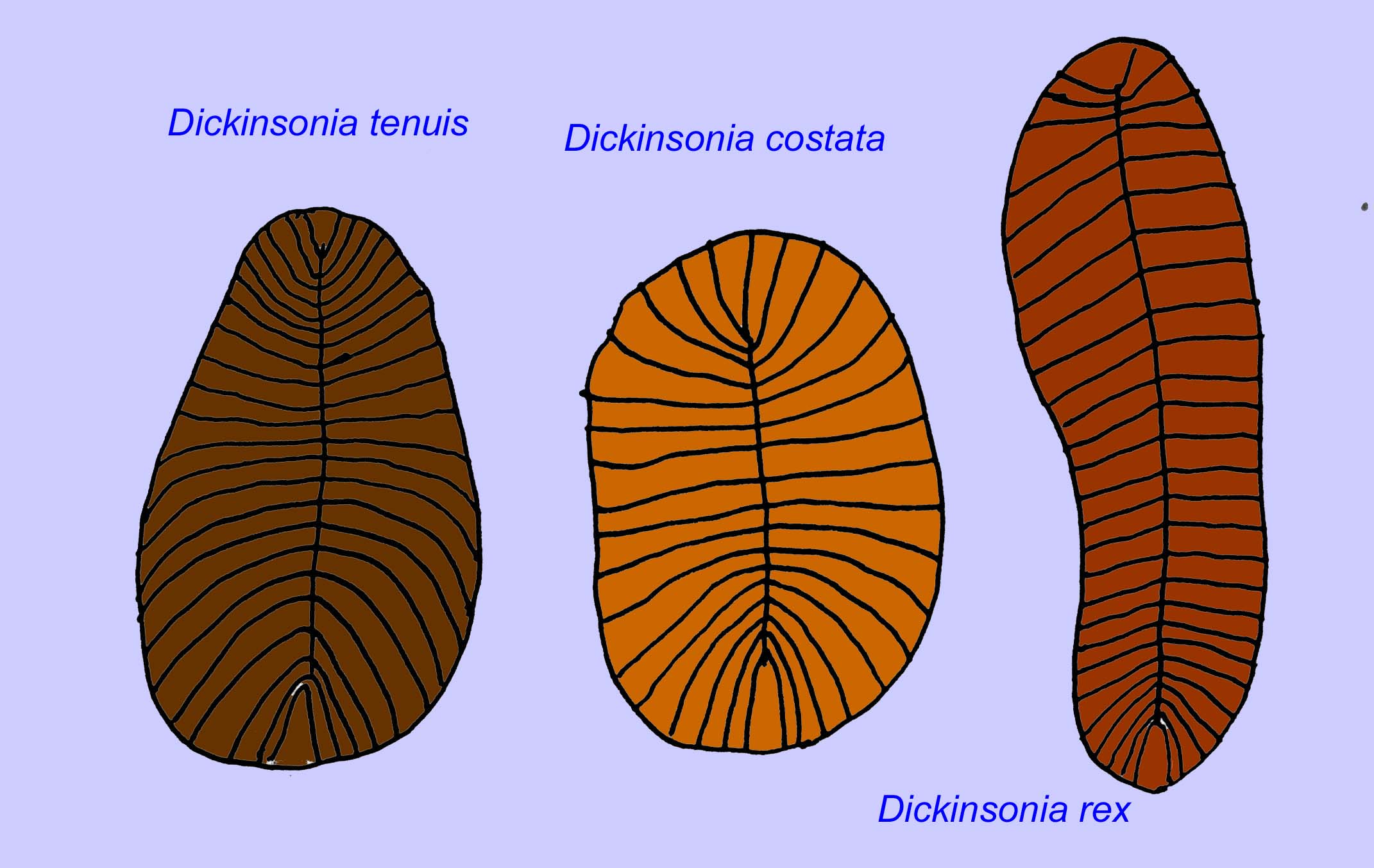
Un certain nombre de vers tels que Paleolina evenkiana et Planolites beverlyensis ont été trouvés à la fin du Protérozoïque et au Cambrien. Les fossiles de Planolites mesurent 1 à 2 mm de large et 5 à 13 mm de long (Bartley, 1998). Les fossiles non segmentés tels que Protechirus, Platypholina, et Vladimissa ne sont pas bien compris. Certains fossiles sont segmentés comme Dickinsonia qui est classé comme un ver annélide. Les plus petits spécimens de Dickinsonia mesurent 4 mm de diamètre tandis que les plus grands mesurent 1 mètre de long. Malgré la longueur de ces vers, ils étaient très fins, moins de 3 mm. Des preuves suggèrent que la quantité d’oxygène dans l’atmosphère à la fin du Précambrien n’était que d’environ un dixième de sa concentration actuelle et l’extrême finesse (et donc la grande surface) de ces vers peut avoir été une adaptation pour la diffusion de l’oxygène. Ces vers étaient communs et répandus. Ils se nourrissaient de débris au fond de l’océan, étant donné les sédiments qui remplissaient souvent leur intestin (Runnegar, 1982). La nature simple de Dickinsonia peut indiquer qu’il s’agit d’un cnidaire plutôt que d’un bilatérien (Brasier, 2008).
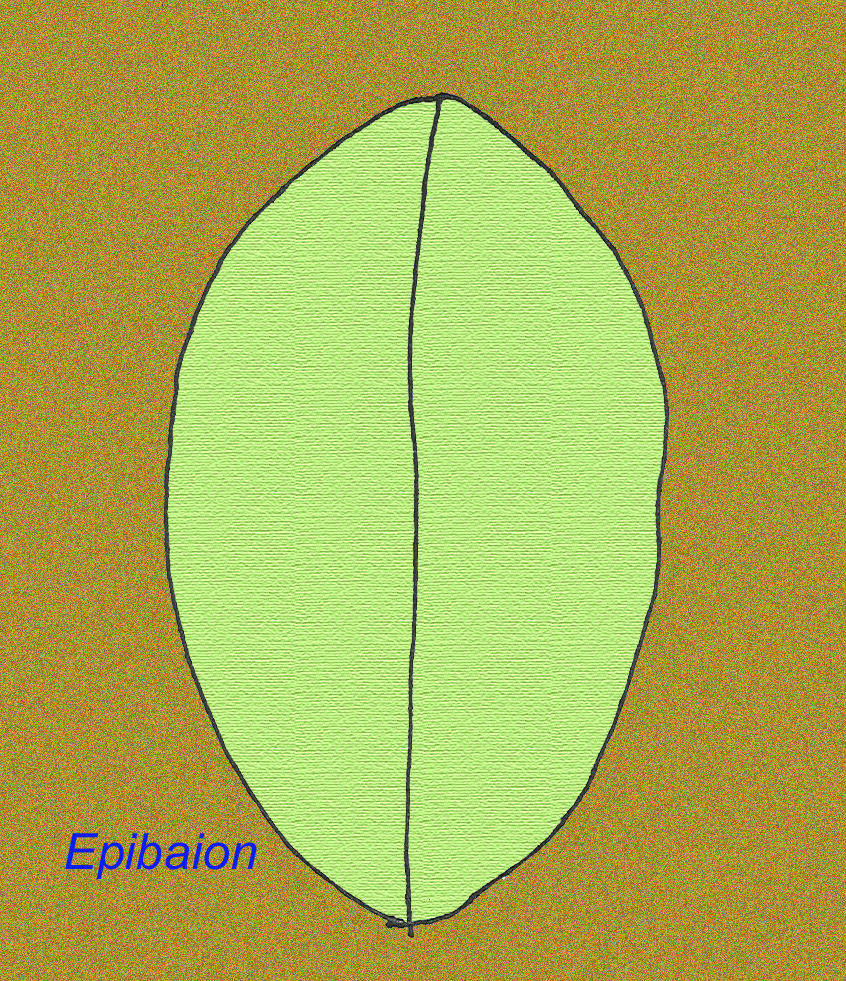
Un grand animal édiacran nommé Epibaion axiferus a laissé des empreintes s’étendant jusqu’à 4,3 mètres sur 0,15 mètre (avec un deuxième spécimen mesurant 2,5 mètres sur 0,35 mètre) (Ivantsov, 2002).


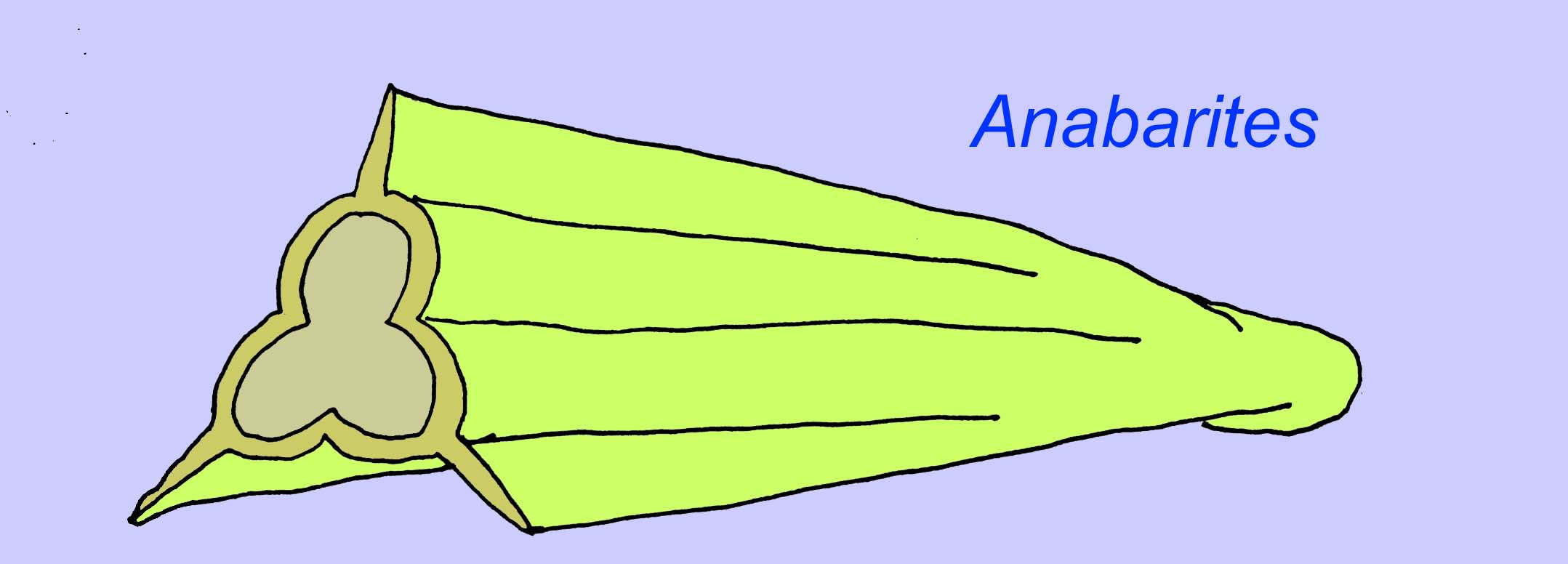
Les anabaritides sont des fossiles à coquille du Cambrien précoce. Certains, comme les anabarites, peuvent remonter à la fin du Précambrien (McMenamin, 1990).
Des traces fossiles de terriers de vers ont été trouvées dans un certain nombre de sites protérozoïques. Ces terriers ont été réalisés par des vers plus évolués que les vers plats. (Bartley, 1998, Narbonne, 1997, Hagadorn, 1998, McMenamin, 1996). Certains cnidaires modernes peuvent former des terriers simples et il est possible que certaines traces fossiles de terriers aient été formées par des cnidaires plutôt que par des bilatériens (Jensen, 2005). Certains terriers de vers sont complets avec des excréments, ce qui implique un système digestif comme celui des vers plus avancés (Morris, 1993a ; Valentine 1994). Des fossiles d’embryons de vers ont été retrouvés depuis le Cambrien le plus ancien (Bengston, 1997).
Y a-t-il des animaux plus évolués que les vers parmi les Édicariens ?
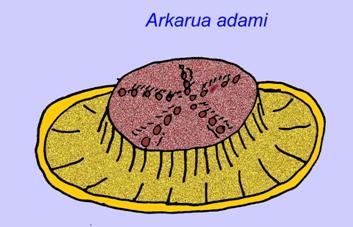
Certaines analyses génétiques ont suggéré que la séparation des principaux groupes d’animaux coelomates s’est produite avant le Cambrien (Gu, 1998). Un certain nombre de fossiles de l’Édiacarien ont été classés dans des groupes de coelomates, apportant un soutien à cette conclusion. Arkurua présente des similitudes avec les échinodermes avec sa symétrie à 5 branches et ses similitudes avec les échinodermes primitifs du Cambrien (les edrioasteroïdes). Les 5 structures sont peut-être des bras d’alimentation menant à une rainure centrale (Gehling, 1987). Deux genres (Redkinia et Spriggina) pourraient être proches de la base des lignées d’arthropodes. Tribrachidium possédait trois appendices sur une de ses surfaces a été trouvé en Australie et en Europe (Wright, 1997b ; McCall, 2006). Certaines espèces de la faune édiacarienne avaient un plan corporel triradial (Tribrachidium, Ventagyrus), tétraradial (Conomedusies) et octaradial (Eoandromeda). Certains les ont interprétés comme des lignées éteintes de cnidaires (Xiao, 2009).
Le fossile édiacarien Ausia a été classé comme un urochordé (Xiao, 2009).
Deux espèces, Phylozoon, et Kimberella (voir ci-dessous), pourraient être des parents de vers segmentés (annélides) ou de mollusques. Kimberella présente une symétrie bilatérale et un possible intestin (Fedonkin, 1997). Kimberella semble avoir été un mollusque qui vivait comme un prédateur marin à nage rapide. Les trous percés dans les coquilles de Cloudina ont été attribués à des prédateurs mollusques édicariens (McCall, 2006). Des fossiles d’embryons de la fin du Précambrien formaient des corps polaires semblables à de nombreux mollusques (Chen, 2006).
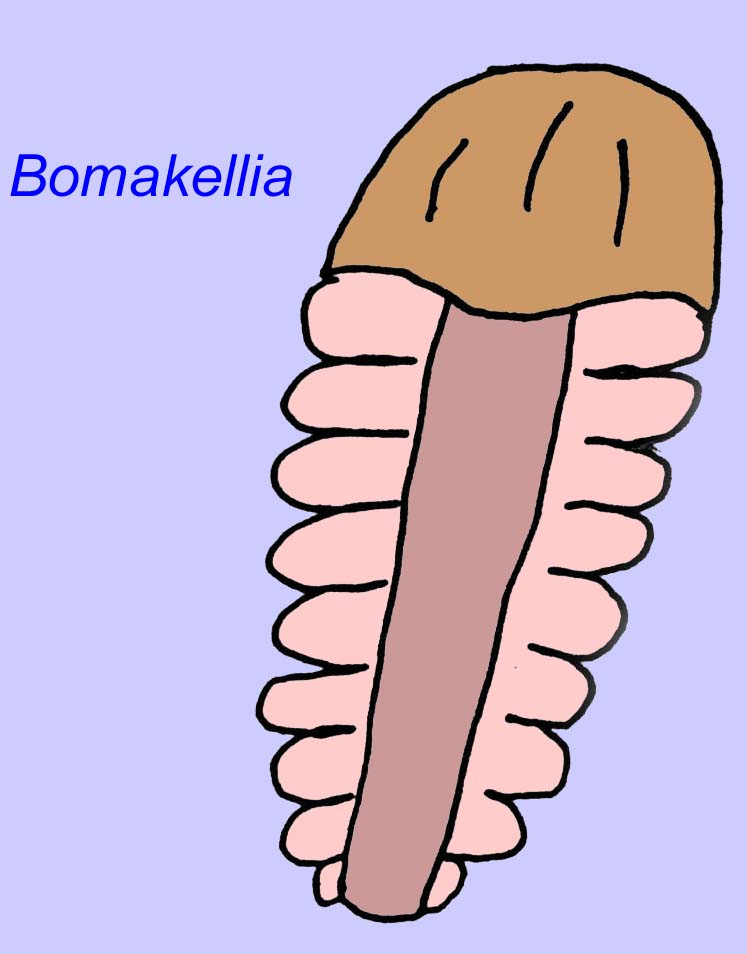
D’autres fossiles peuvent être liés à des arthropodes, dont certains (comme Bomakellia) ont une structure qui a été comparée à la tête-bouclier des trilobites (un type commun d’arthropode de l’ère paléozoïque ; McCall, 2006).
Certaine faune de l’Édiacarien avait un plan corporel triradial (Tribrachidium, Ventagyrus), tétraradial (Conomedusies) et octaradial (Eoandromeda). Certains les ont interprétés comme des lignées éteintes de cnidaires (Xiao, 2009).
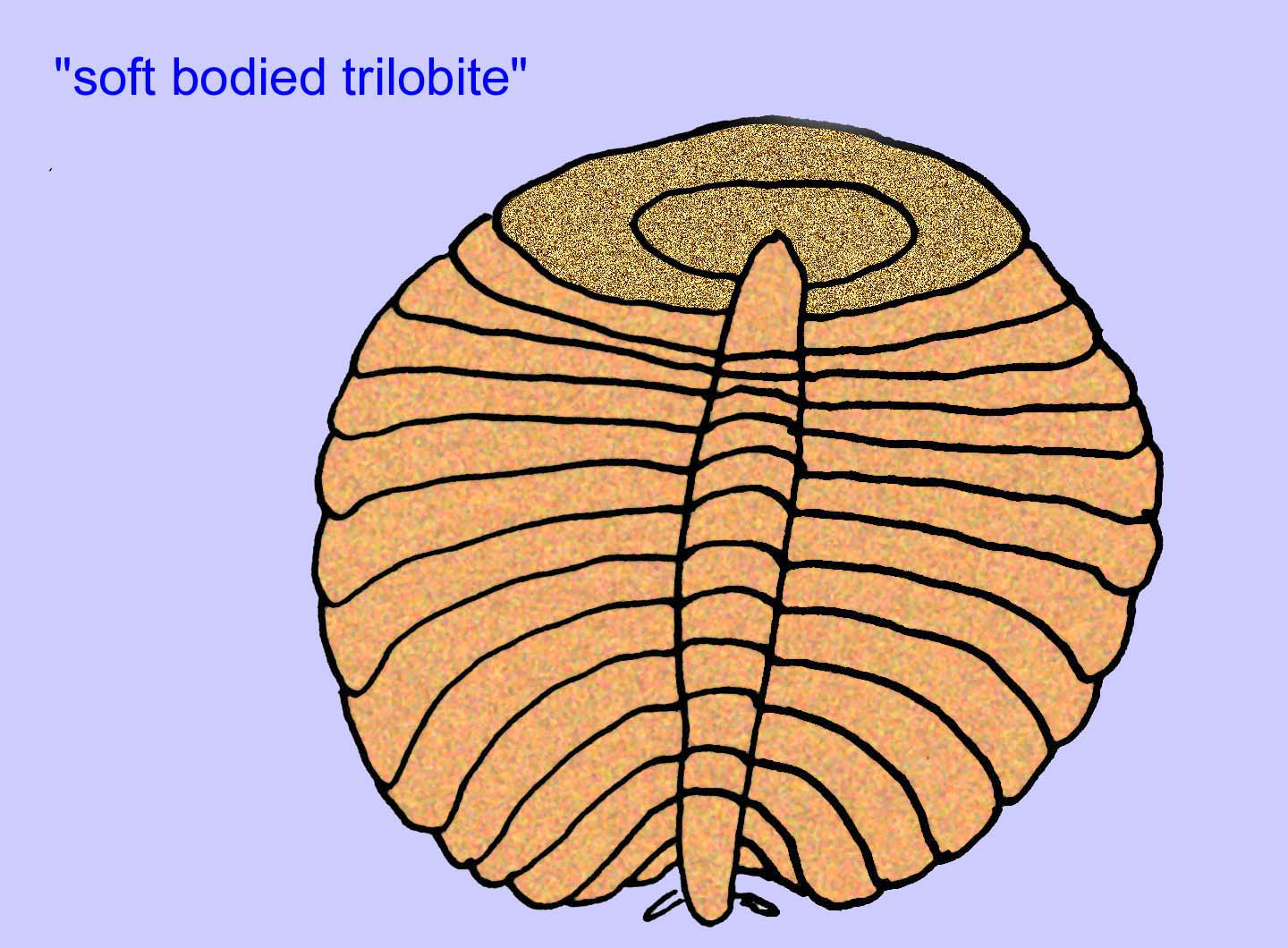
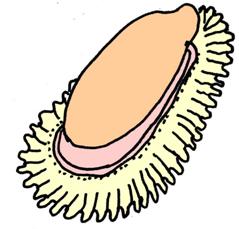

.