最初の動物
最も原始的な生物は単細胞で、生物全体が1つの細胞で構成されています。 生物の中には、個々の細胞として生きるものや、細胞の集まりであるコロニーとして生きるものがあります。 コロニーの細胞が分離しても、個々の細胞として生き続けることもできるし、新しいコロニーを形成することもできる。 特定の仕事に特化した細胞はない。 現代の藻類であるVolvoxがその良い例で、隣の写真はVolvoxのコロニーです。
植物、紅藻類、褐藻類、菌類、動物などでは、数十回にわたって多細胞化が独自に進化しています。
多細胞化は、植物、紅藻類、褐藻類、菌類、動物などで数十回、独自に進化してきました。例えば、紅藻類では、単細胞種(Chlamydomonas)、一般的な4細胞種(Basichlamys)、50,000細胞のコロニーを形成する種(Volvox)などがあります(Sachs, 2008)。
多くの藻類はこのような単純なコロニーで生活していますが、実際には異なる仕事をするために特化した細胞を持っている藻類もあります(根のような機能と葉のような機能を持つ組織を持っている)。
藻類の最初の化石は約21億年前から知られていますが、約10億年前になると、知られている藻類の種の多様性が増してきます。 真核生物である藻類は、前生代末期の氷河期を経て、6億年前頃に突然多様化したようです(Cavalier-Smith, 2000)。
5億7000万年前には、様々な解剖学的・生殖学的特徴を持つ藻類が知られています(Xiao, 1998)。 化石記録には、以下に描かれているような多様な先史時代の藻類が保存されています。
菌類
エディアカラ以前の化石の多くは、バクテリアの単純な多細胞コロニー、緑藻類や紅藻類を含む真核藻類、そして菌類に似た形態を表しています。 (Butterfield, 2009; Xiao, 2009)。
1億年前の琥珀では、肉食性の菌類が菌糸環を形成して線虫を捕食する様子が観察されました(Schmidt, 2007)。
最初の動物たち
エディアカラ動物群は、最初に発見されたオーストラリアのエディアカラ丘陵にちなんで名付けられた生物群です。 エディアカラ動物群からは、270種以上からなる数千個の標本が報告されており、まだ記述されていないものも多い(Fedonkin, 1990; Minnerop, 2000; Waggoner, 1998; Shen, 2008)。 最初のエディアカラの動物は1872年に記載され、その後1908-14年に2回目の発見がありました(McCall, 2006)。
- THEY ARE OLD
12億年以上前の化石層には動物が含まれています。放射状に柔らかい体を持つ生物の化石が含まれているだけでなく、粘液を出すミミズが作ったと思われる巣穴も含まれています(Rasmussen, 2002; McCall, 2006)。 インドの遺跡では、ミミズの巣穴の年代が11億年前とされていますが、この年代に異論もあります。 最近では、原始的な動物(クラゲの親戚)やミミズの巣穴の痕跡が見られる別の遺跡の年代が、最も若いもので12億年前とされています(Seilacher, 1998; Kerr, 1998b; Morris, 1993)。 5億8千万年前の先カンブリア紀の地層には小型の二枚貝の化石があり、5億5千5百万年前以降には大型の二枚貝が存在する(Condon, 2005)。 エディアカラン期は、マリノン・スノーボール・アース氷河の後の6億3500万年前と定義されています(ただし、5億6200万年前のガスキア氷河はエディアカラン期の中で起きています)(Xiao, 2009)。
現代の生物の分子比較も、カンブリア紀以前に動物がさまざまな系統を生み出していたという結論を裏付けています。 例えば、129個のタンパク質を用いたある研究では、真核生物は9億5000万年前から12億6000万年前の間に、動物は7億6100万年前から9億5700万年前の間に、そして腔腸動物の系統は6億4200万年前から7億6100万年前の間に分離したことが示唆されています(Douzery, 2004)。 他の分析でも同様の結果が得られている(Peterson, 2004)。また、分子時計研究による真獣類(刺胞動物と二枚貝)の起源の分子推定値は、5億7000万年前から11億年前まで様々である。 二枚貝類の起源は5億8000万年前と推定されている(Peterson, 2005)。 オーストラリア、インド、テキサスで発見された動物の化石の年代は10億年以上前、ロシアで発見された分節動物の年代は8億9000万年から10億年前とされている(De, 2005)。 先カンブリア時代の動物のエキサイティングな発見のため、地質学的時間スケールに新しい時代が追加された。これは、1891年(炭素紀が分割されたとき)以来、層序的に新しい時代が定義されたことを意味する。 エディアカラ期とは、6億3000万年前からカンブリア紀の始まりである約5億4200万年前までの期間と定義されています(Knoll, 2004)。
これらの年代にはどのような意味があるのでしょうか。 つい最近まで、約5億4500万年前に始まった「カンブリア紀の爆発」で、多様で複雑な動物が突然出現したと考えられていました。 しかし現在では、カンブリア紀の少なくとも6,000万年前から、あるいは6億年前から動物が存在していたことがわかっています。 6,000万年というと、先カンブリア時代の広大な時間を考えると、それほど長い時間ではないと思われるかもしれませんが、その間に動物の進化は大きく進んでいます。 魚類が両生類に進化するのに約6,000万年、両生類が爬虫類をはじめとするさまざまな系統に多様化するのに約6,000万年かかっている。 二畳紀の終わりに始まる6千万年の期間には、2つの大量絶滅が起こる。 この時期は、シナプシッド型爬虫類と原始的なアーコサウルスが支配する世界から始まり、恐竜、翼竜、海棲爬虫類などの多様な系統が存在するジュラ紀に終わる。 そして約6千万年後、恐竜の絶滅を生き延びた原始的な哺乳類が放射状に広がり、新生代の多様な化石・生物群を形成したのです。


2) THEY ARE SIMPLE ANIMALS
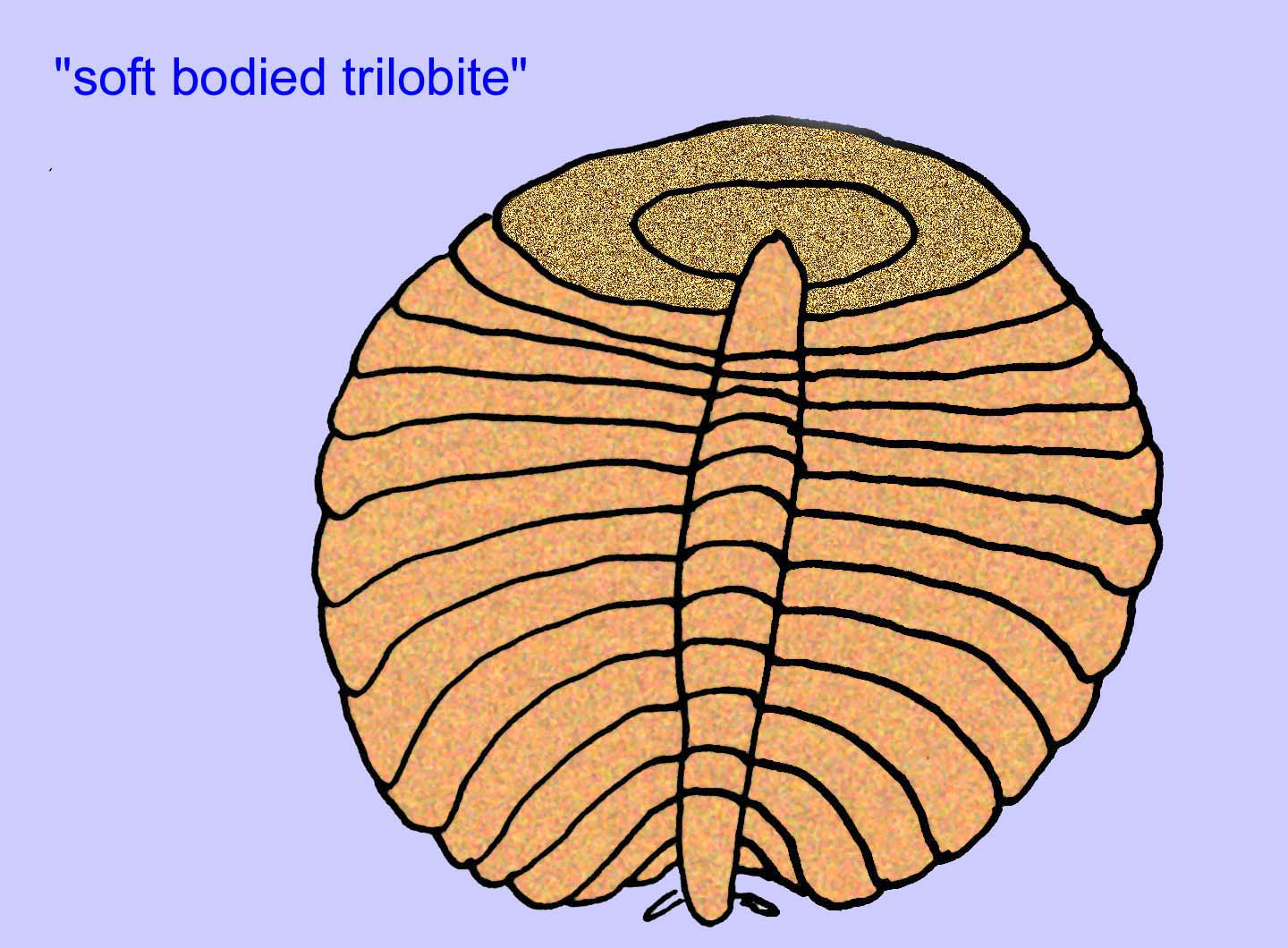
エディアカラの生物のほとんどは1インチ以下の大きさですが、中には数フィートの長さのものもあります。 彼らは奇妙で、ほとんどが現在生きているものとは全く異なります(多くの人は、彼らをVendobiontaというグループと呼んでいます)。 ほとんどの生物には、頭や尾、内と外、表と裏、歯、目がない。 ほとんどの生物には、明らかな循環系、神経系、消化器系、内臓器官がない。 循環器系、呼吸器系、消化器系、排泄器系を持たない動物にとっては必要不可欠な条件である。 いくつかのサイトでは、30もの異なる分類を持つ多様な動物がいます(Buss, 1994)。
動物とは何か? 過去には、植物(第一次世界大戦の時点での結論)、非常に大きな単細胞生物、ユニークな地衣類、完全に絶滅した動物、現代動物の祖先を含む動物群など、これらの生物に関する多様な意見がありました。 もともとは、現代の動物群が誕生するずっと前に絶滅したと考えられていたが、現在ではカンブリア紀まで生き残ったグループがあることが明らかになっている。一部はメタゾア以外の動物の化石を表しているかもしれない(Droser, 2006)。 先カンブリア時代の底生動物のバイオマス量は、現代の社会で観察されるバイオマス量に近かった可能性があることを、円盤動物Aspidellaの個体密度が示しています(Gehling, 2000)。
エディアカラ以前の化石の多くは、バクテリア、真核藻類(緑藻、紅藻を含む)、菌類に似た形態などの単純な多細胞コロニーです。
最も単純な動物の一つである海綿は、古生代の海綿に似たエディアカラの堆積物(Palaeophragmodictya属など)から同定されています。 これらの海綿は、直径1.5〜10cmにもなる(Gehling, 1996; Kerr, 1998a)。 また、エディアカラの岩石中にもスポンジの胞子が存在します(Xiao, 2000)。
エディアカラの化石の中には、スポンジよりも原始的な菌類や基底動物と解釈されているものもあります(Xiao, 2009)。
刺胞動物
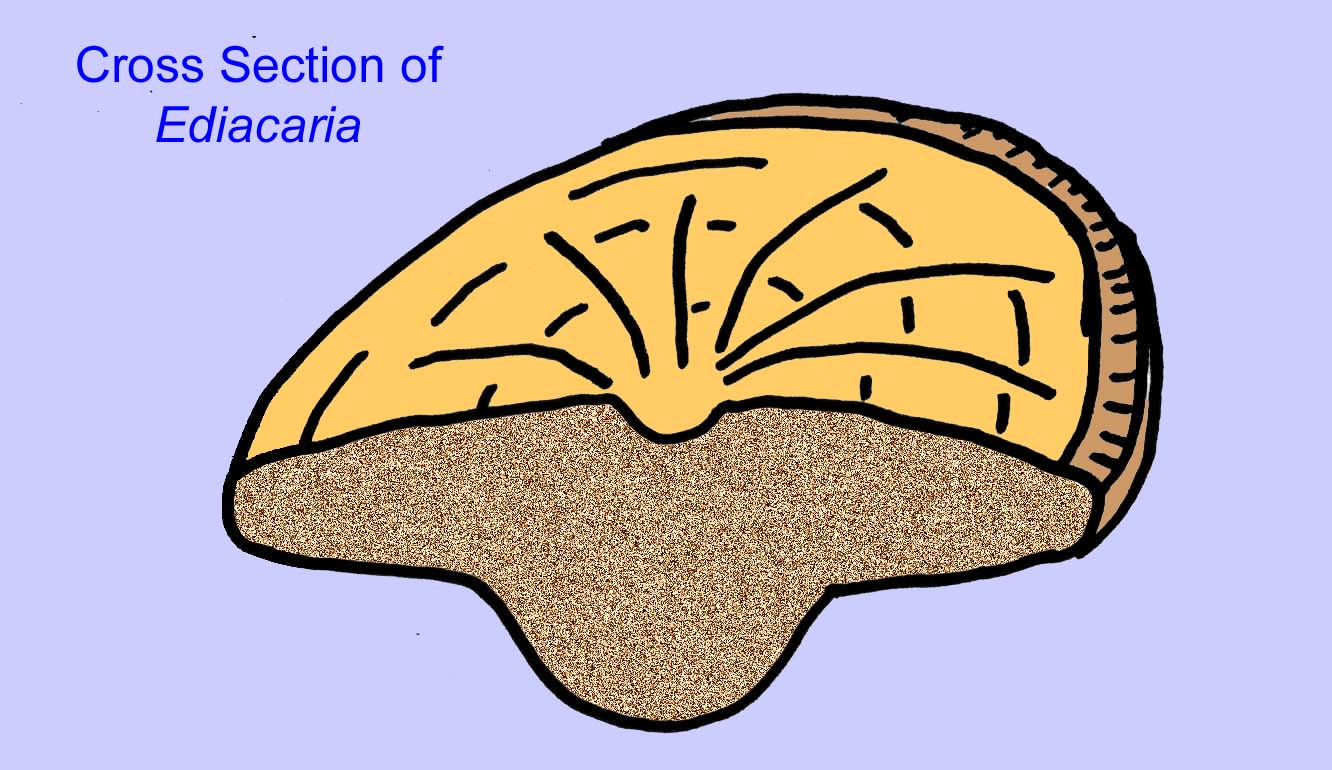
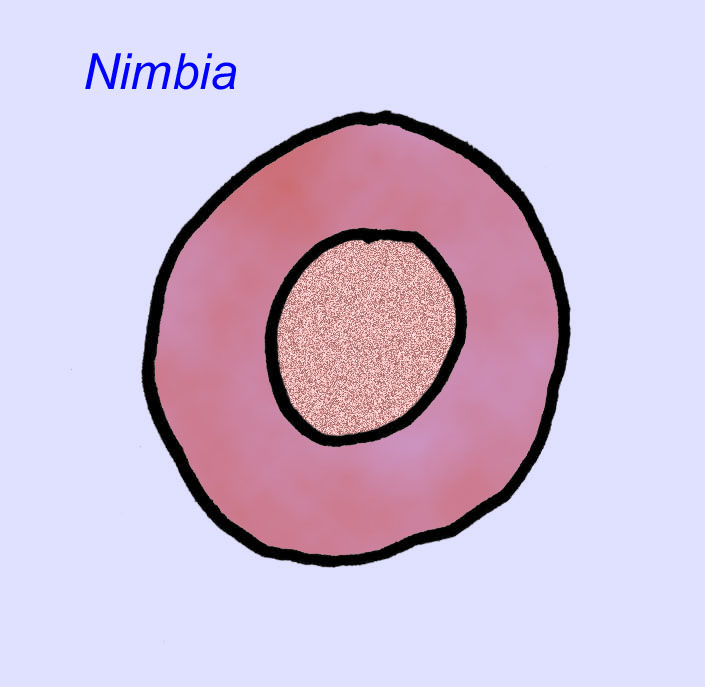
 ivid cnidarian
ivid cnidarian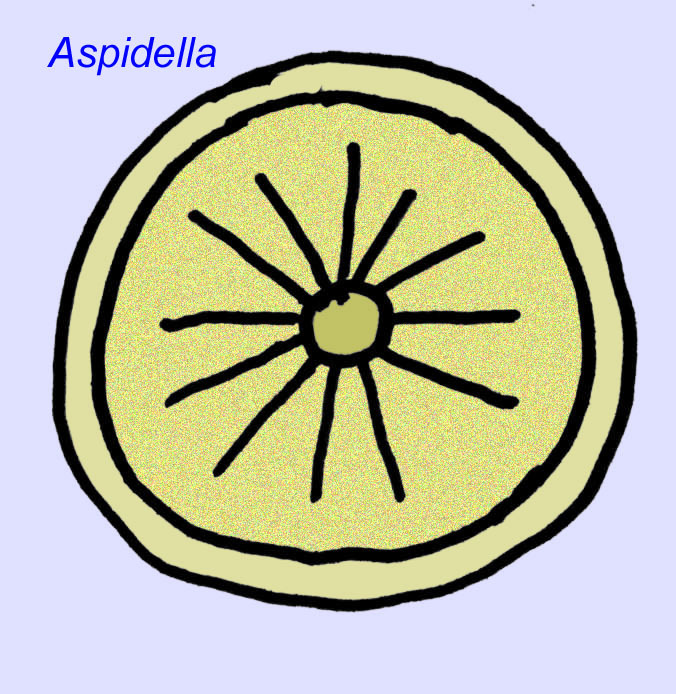
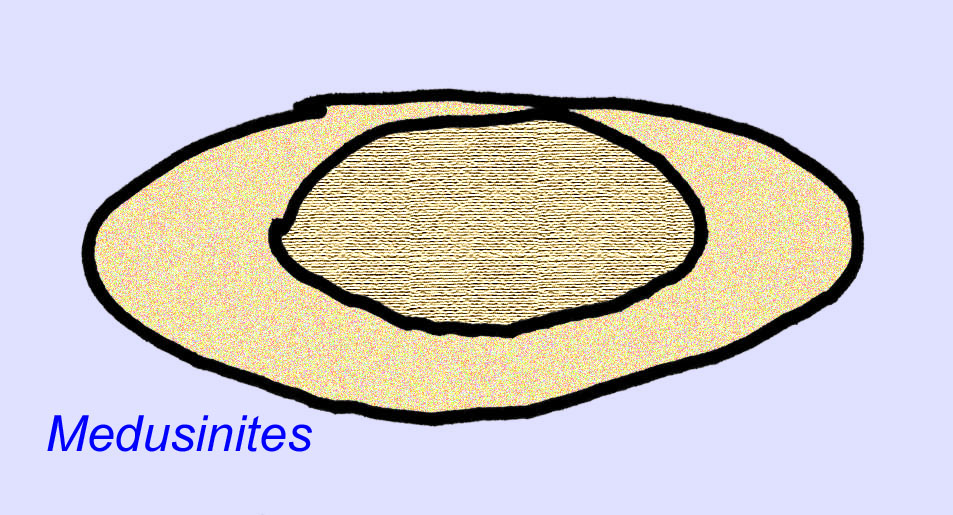
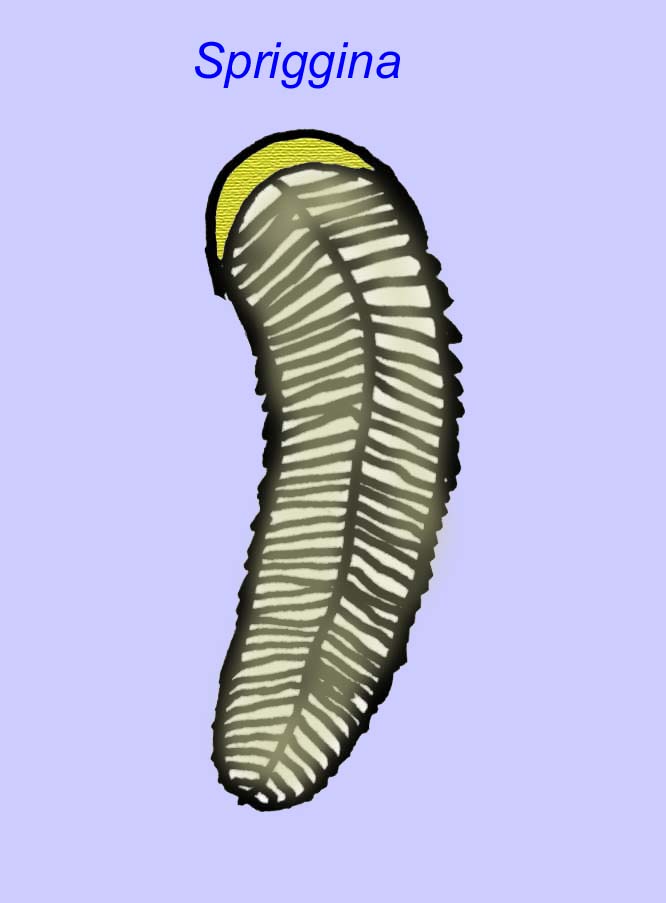
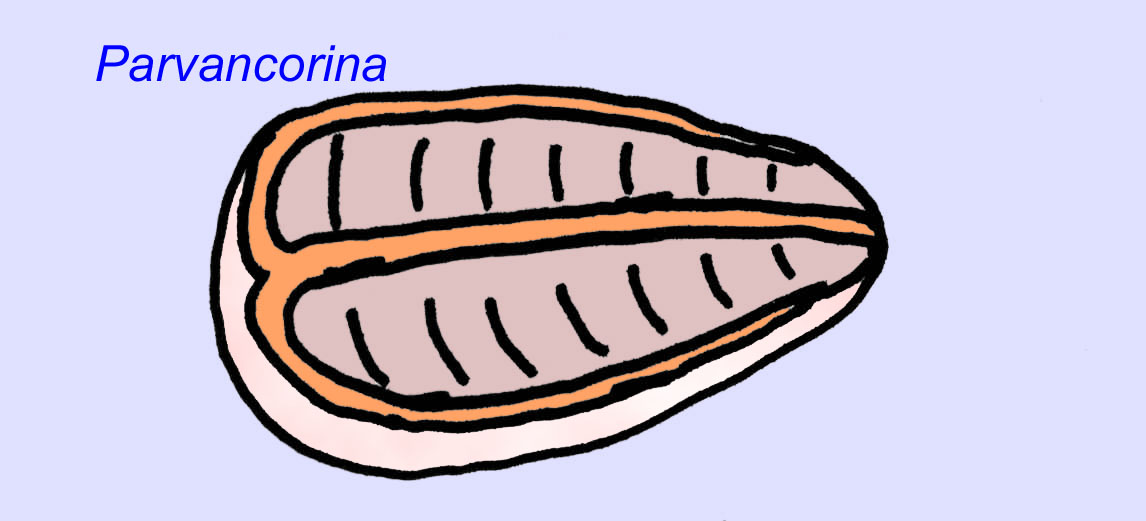
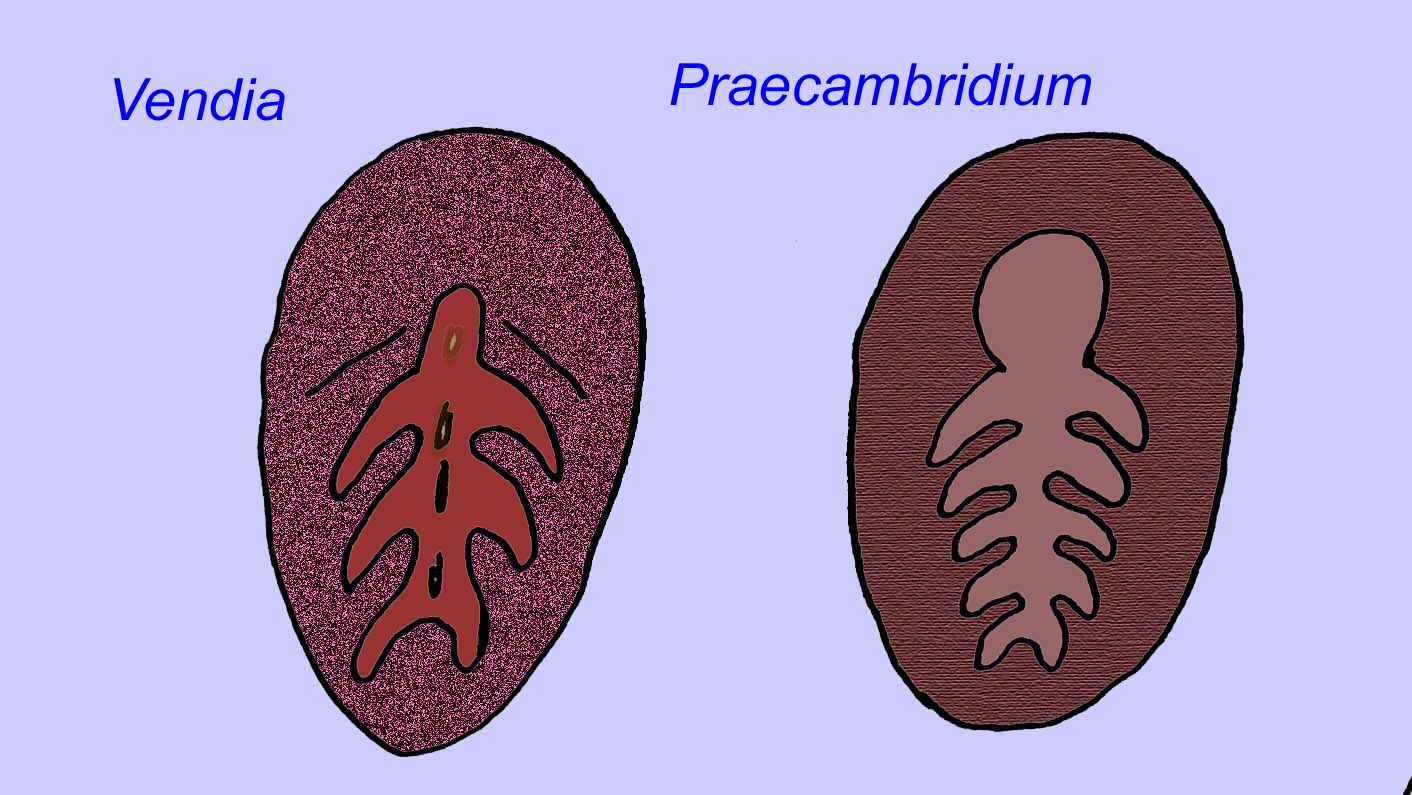
エディアカラの化石の多くは、初期の刺胞動物に分類されるべきだという一般的なコンセンサスがあります。 現代のクラゲ、ウミウシ、ウミウチワなどを含む動物のグループです。 EdiacariaやNimbiaのように、半径方向に対称で表面的には刺胞動物に似ているが、口の形をしていないものもあり、このグループから除外する理由になっているかもしれない。 最初の刺胞動物が口を持っていたかどうかは定かではないが、後の形態はすべてそうである)。 Albumares brunsaeのように、刺胞動物と同様の胃血管系を持つものもある(Lipps, 1992; Sokolov, 1985)。 扁平で丸い体の片側に中央に円盤状の構造を持つものも多い(Crimes, 1995; Bertrand-Sarfati, 1995)。 一部のエディアカラの動物(Spriggina, Parvancorina, Praecambridium)は、上面と下面(後面と前面)に区別がある。 また、エディアカリアのように、放射状の線や周辺部の肥厚を持つものもあった(Crimes, 1995)。 エディアカラの刺胞動物の中には、別の、しかし絶滅したサブクラスとして分類されているものもあります(McCall, 2006)。
これらの動物は鉱物性の骨格を持っていませんでした。
これらの動物は鉱物性の骨格を持っていませんが、ほとんど、あるいはすべての動物(サイクロメドゥーサやエルニエッタなど)は、泥の中で印象を残すことができる何らかの硬い骨格を持っていました。 内部骨格を構成する組織の種類は不明です(Crimes, 1995; Dzik, 1999)。
マッケンツィアも刺胞動物と思われます(そうでなくても、ほぼ間違いなく後生動物です)。 消化腔内のフードボーラスや筋骨格の間接的な証拠があります(Morris, 1993a)。 Sinocyclocusは基底の刺胞動物(あるいは基底の後生動物)である可能性があり、その先端の出芽はいくつかの刺胞動物(タブレイト・コーラルなど)のそれに似ている(Xiao, 2000)。
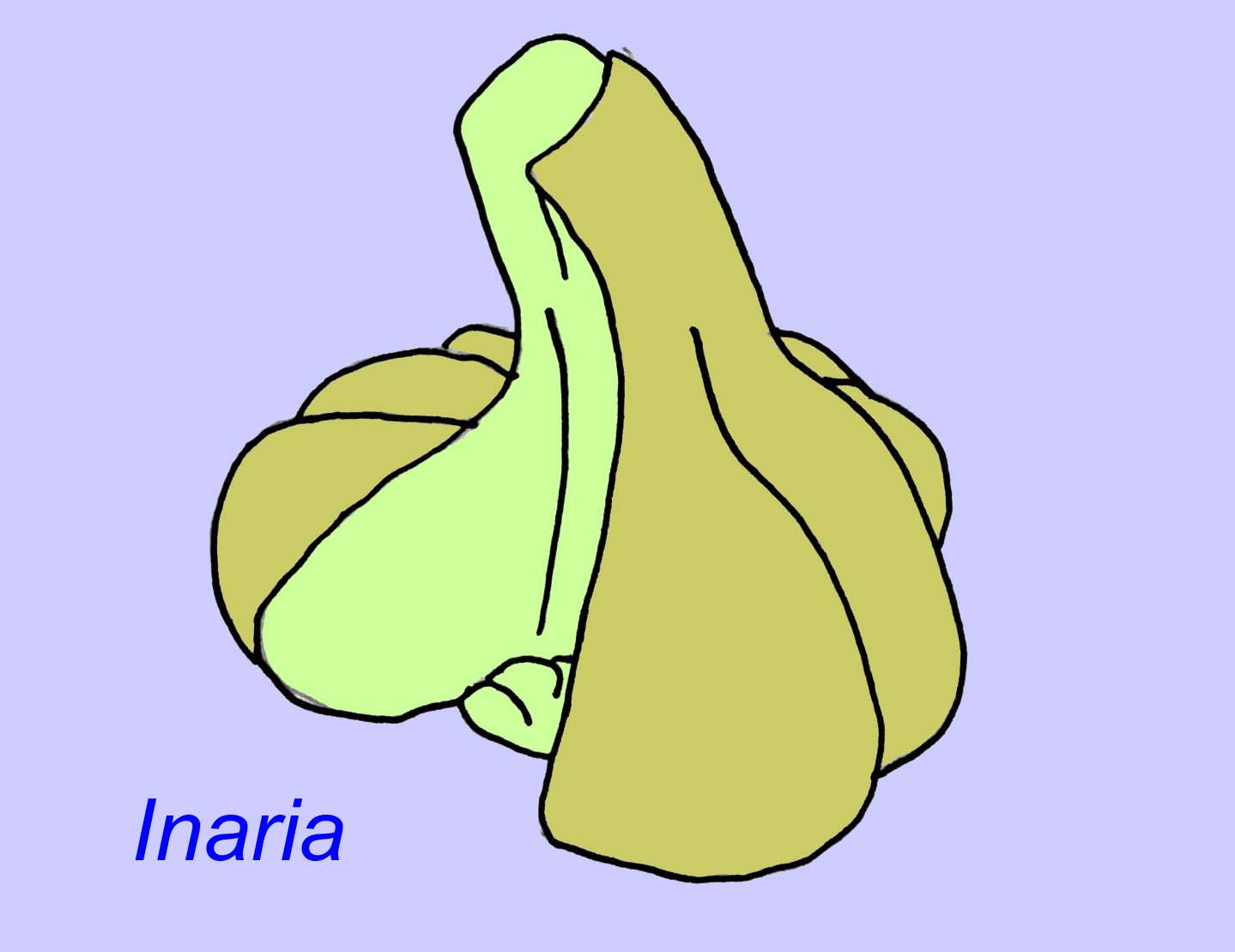
イナリアは、口縁部、咽頭部、そして現代の多くのサンゴに見られるような光合成を行う生物を収容する部屋を持つ、刺胞動物の一種です(McMenaman, 1990, Gehling, 1988)。
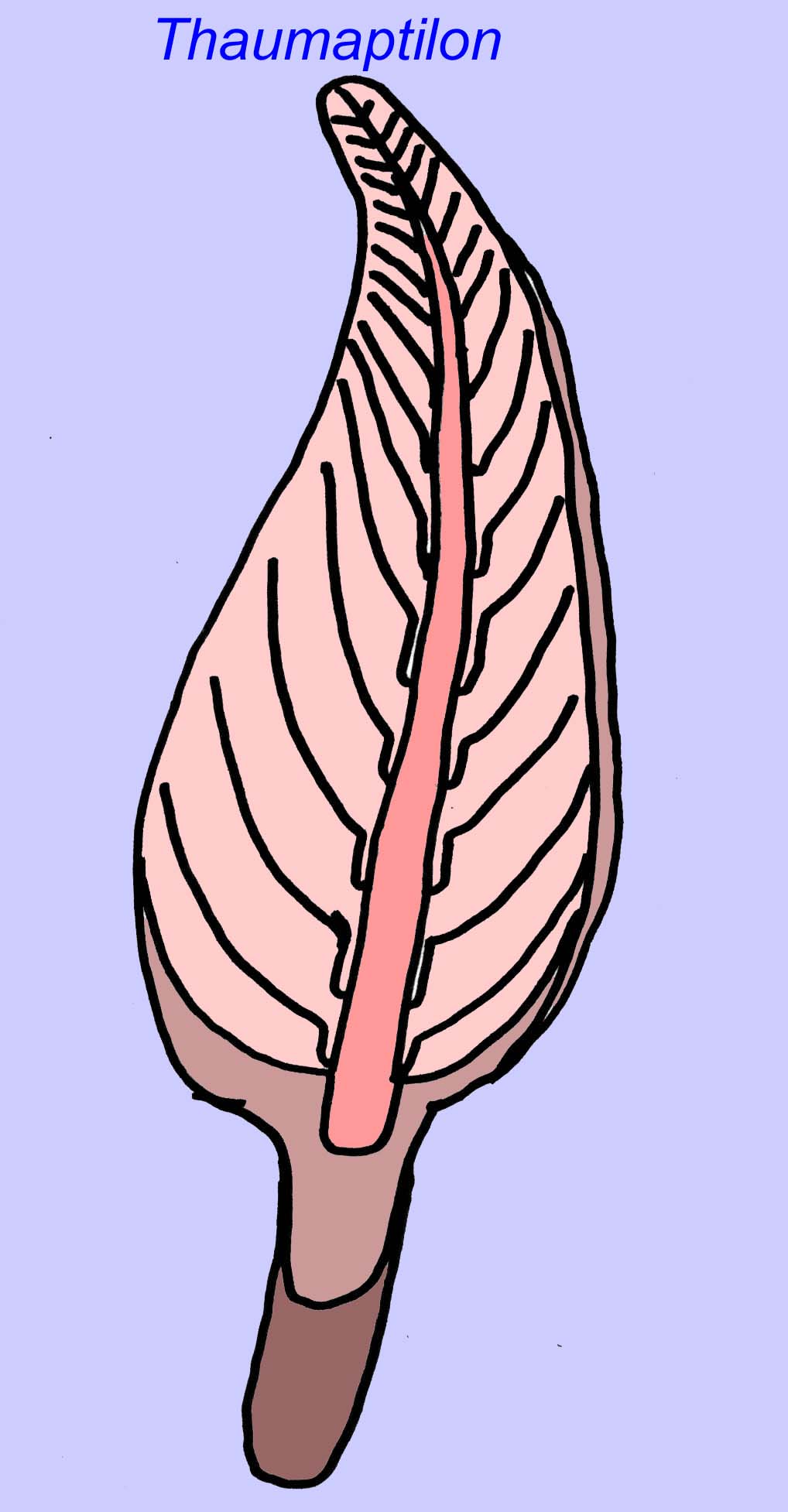
Thaumaptilonは、現代のウミガラスに関連していると思われる、前足のような動物でした。
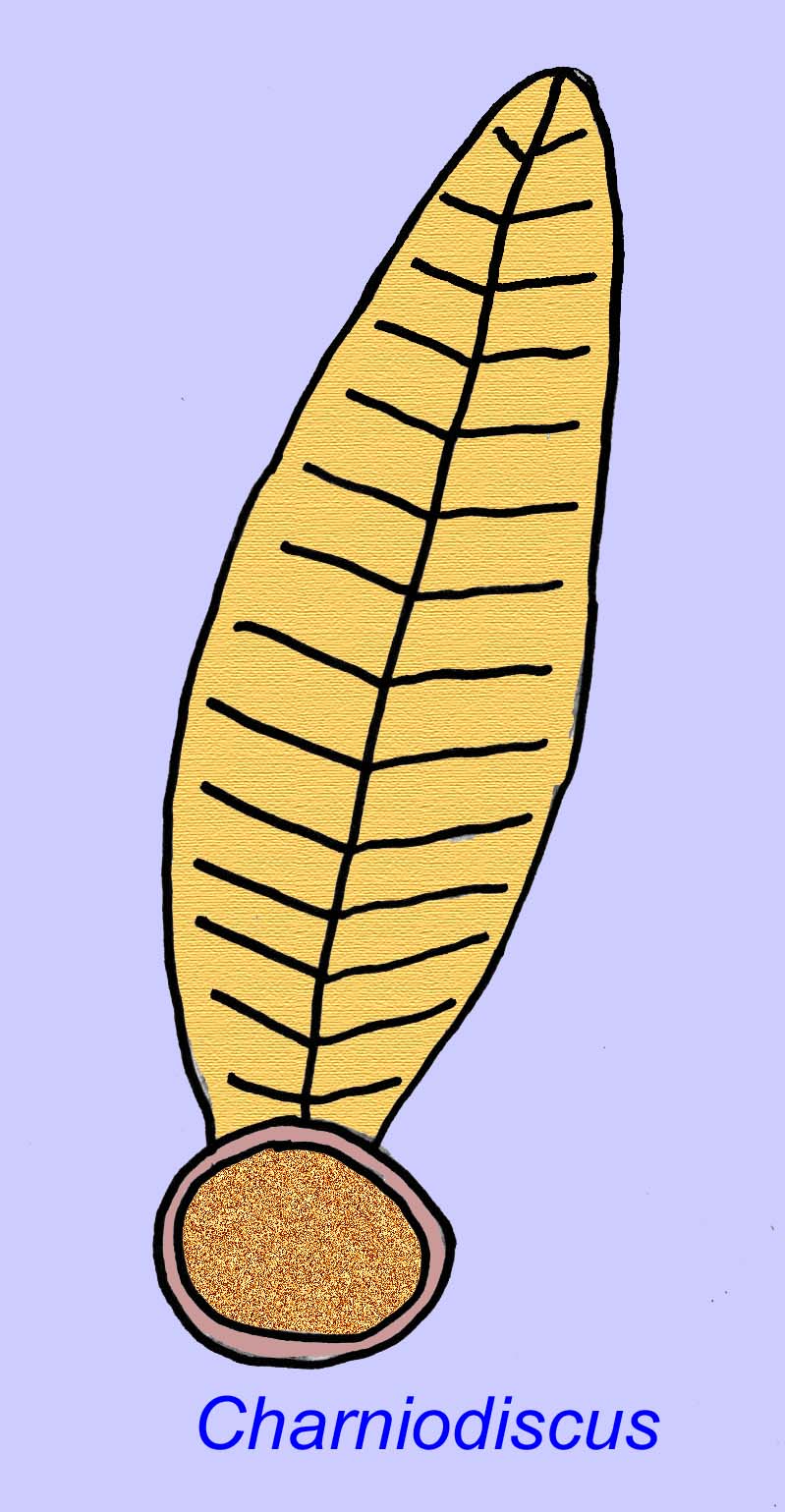
Thaumaptilonは、現代のシーペンに関連すると思われるフロントのような動物で、粒子状の破片を食べ、底に固定するためにホールドファストを使っていたと思われます(Morris, 1993a; 1993b)。 Charniodiscusのフローンは1.2メートルの長さにもなります(Sokolov, 1990)。
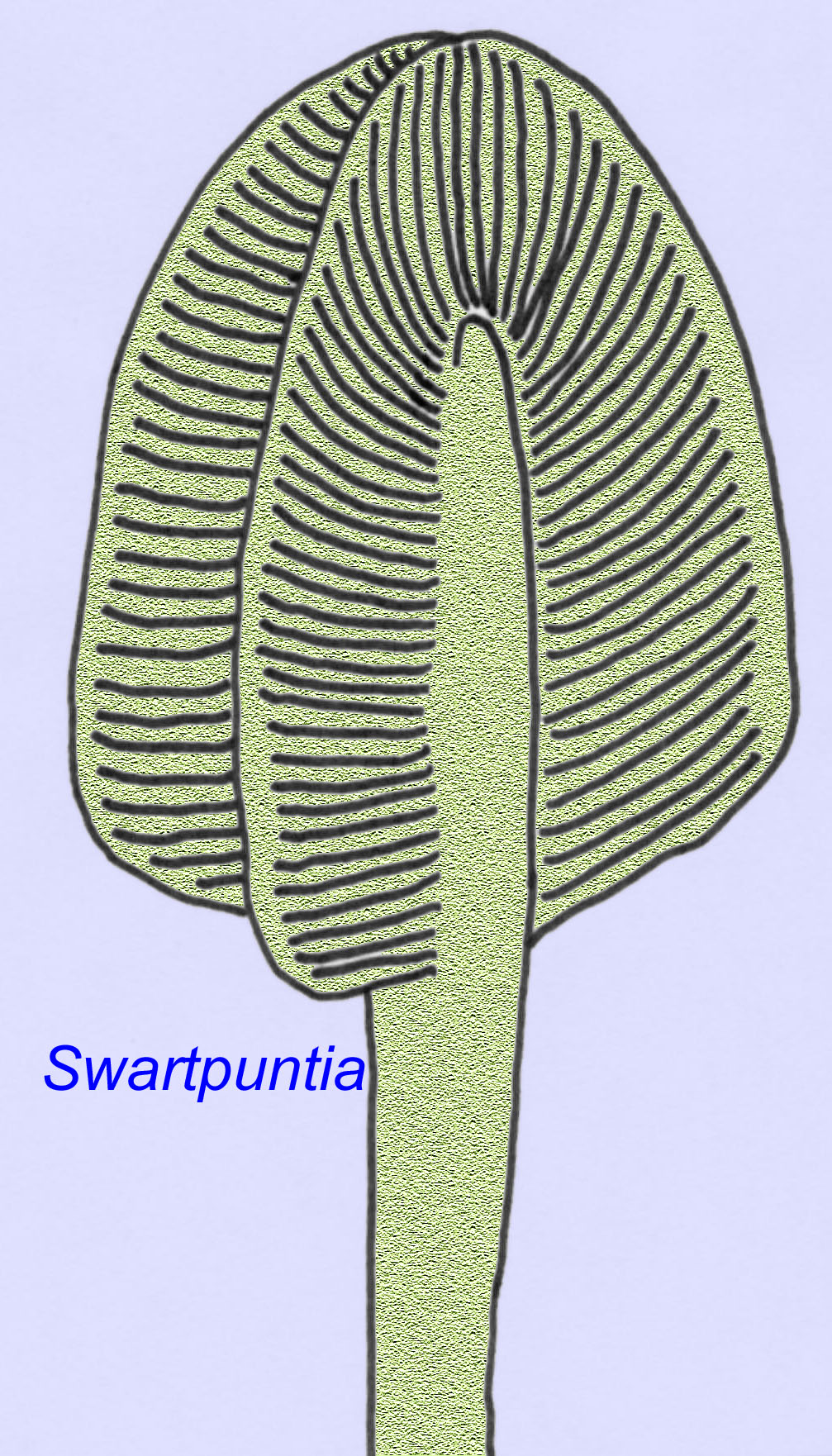
多くのエディアカラの動物(Pteridinium, Swartpuntia, Charnia, Charniodiscusなど)は、直立した茎にセグメント化された花びらが結合しています。
Charnodiscusの1つの標本の長さは1.2メートルでした。
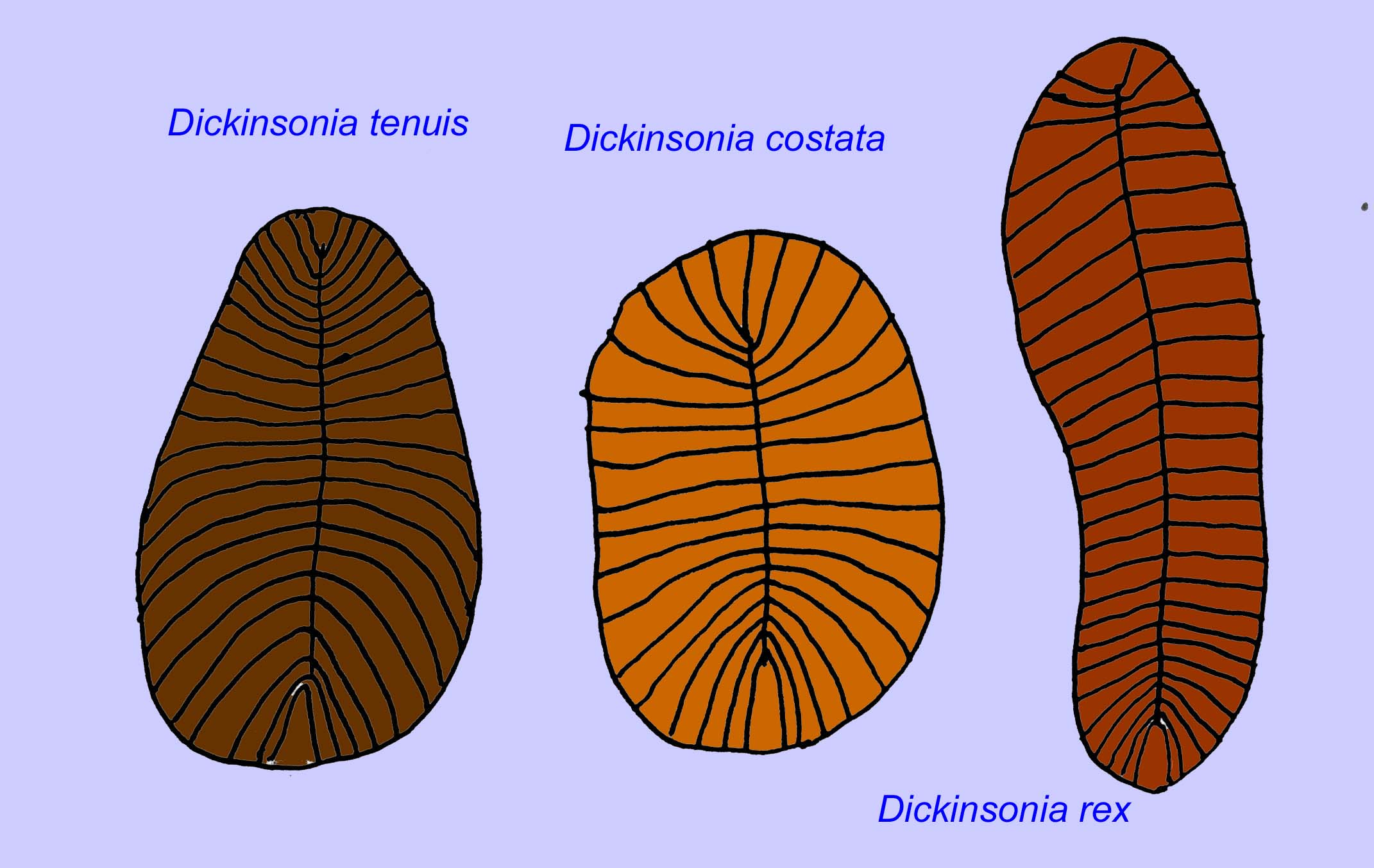
Swartpuntiaの顕微鏡解剖は、初期の虫であるDickinsoniaと非常によく似ています(Narbonne, 1997; McCall, 2006)。 また、5億7千万年前の二枚貝と思われる胚の化石が知られています(Xiao, 1998)。

WORMS
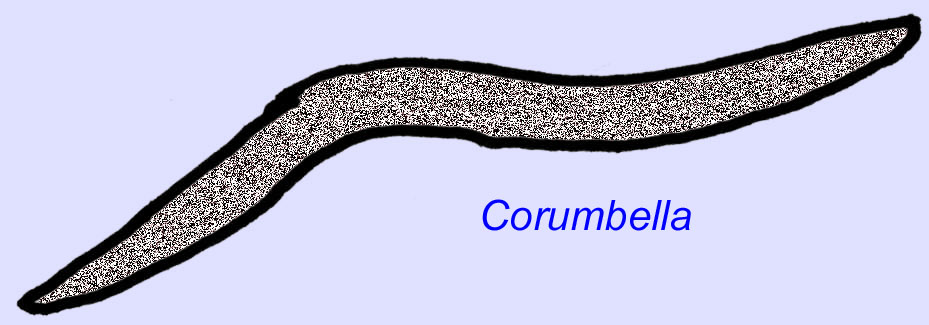
エディアカラのワームはアジアから知られています。 アジア、ヨーロッパ、アフリカ、オーストラリア、北米から知られています(McCall, 2006)。 Archaeichnium, Cloudina, Corumbella, Onuphionellaなど多くのエディアカラの動物は管状でした。 オヌフィオネラは環形動物の虫と比較されるが、コロンベラは刺胞動物のポリプが作るチューブと比較される。 Archaeichniumが柔軟であるのに対し、Cloudinaは鉱物化した骨格に覆われていた。 クラウディナのチューブ(最大長4cm以下)には環状のものと滑らかなものがあるが、これは保存状態の違いによるものかもしれない(Hagadorn, 2000)。 クラウディナは鉱物化した骨格を持つ最初の動物として知られており、標本の3%には捕食を示す穴が開いている。 この捕食が、カンブリア紀の動物の多くに見られる硬い構造を促したのかもしれない(Bengston, 1992)。 Cloudinaは南アメリカ、ヨーロッパ、中東、東南アジアの先カンブリア時代から知られており、親戚は北アメリカの初期カンブリア時代(三葉虫が普及する前)から知られている(Grant, 1990)。 Cloudinaの化石は、カンブリア紀が始まる少なくとも600万年前から知られています(Condon, 2005)。
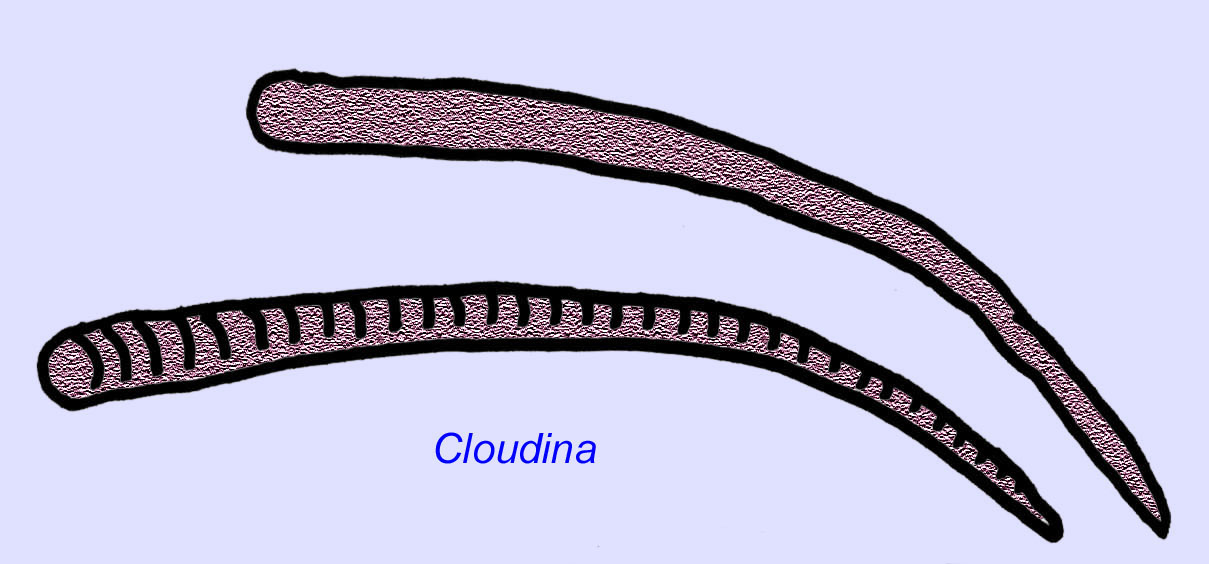
認識されている2種のCloudinaの他にも、多くの化石種が先カンブリア時代後期に石灰化した殻を保有していました(McCall, 2006)。 Cloudina、Namacalanthus、Conotubusは、カンブリア紀初期にも知られている先カンブリア紀の骨格を持つ動物です(Shu, 2008)。
Paleolina evenkianaやPlanolites beverlyensisなどのミミズが、原生代末期からカンブリア紀にかけて多数発見されています。 プラノライトの化石は、幅1〜2mm、長さ5〜13mmである(Bartley, 1998)。 Protechirus、Platypholina、Vladimissaのようなセグメント化されていない化石はよくわかっていない。 環形動物に分類されるDickinsoniaのようにセグメント化された化石もある。 ディキンソニアは、小さいものでは直径4mm、大きいものでは1mにもなります。 このような長さにもかかわらず、3mm以下の非常に細い虫でした。 先カンブリア時代末期の大気中の酸素量は、現在の10分の1程度であったことが示唆されており、これらの虫の極端な薄さ(つまり表面積の大きさ)は、酸素拡散のための適応であったと考えられる。 これらのミミズは一般的で広範囲に生息していました。 彼らは海底の瓦礫を食べていたので、しばしば腸を満たしていた堆積物を与えられていました(Runnegar, 1982)。
Epibaion axiferusという大型のエディアックラン動物は、4.3メートル×0.15メートル(2.5メートル×0.35メートルの第2標本もある)の印象を残しています(Ivantsov, 2002)。


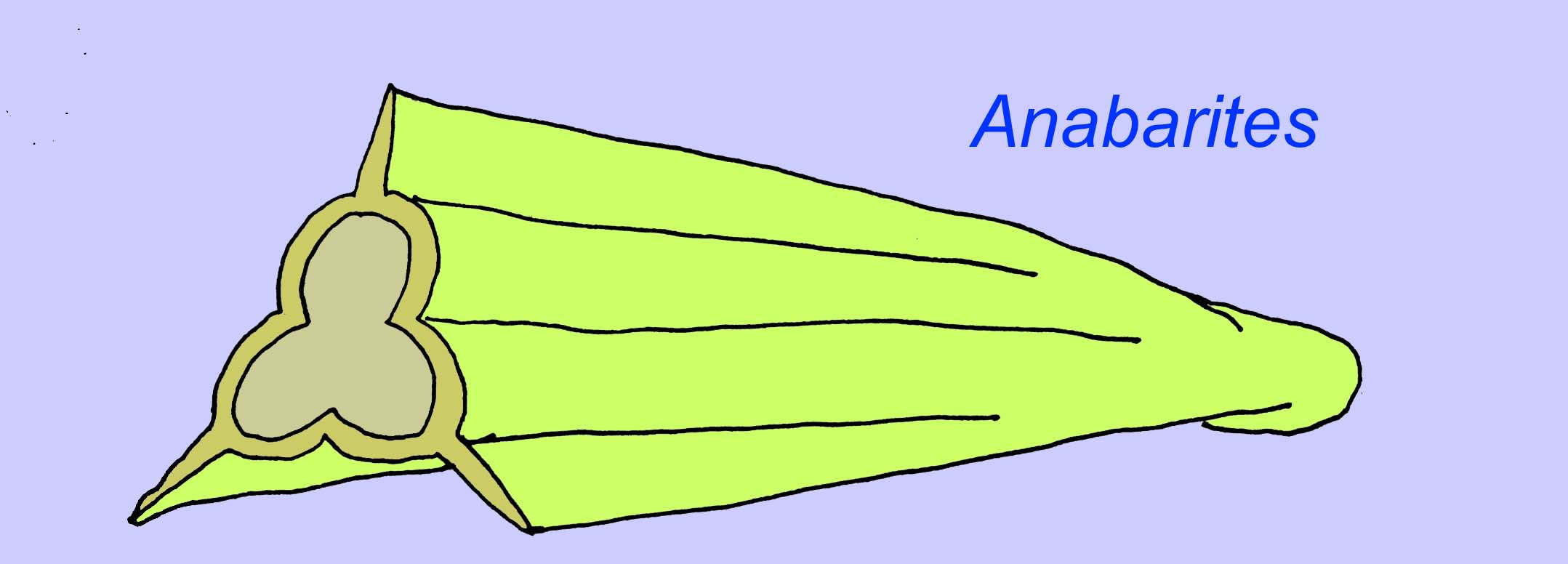
アナバリット類はカンブリア紀初期の殻付きの化石です。 Anabaritesのように、先カンブリア時代の終わりまでさかのぼるものもあります(McMenamin, 1990)。
原生代の多くの場所で、ミミズの巣穴の痕跡化石が発見されています。 これらの穴はヒラムシよりも進化したミミズが作ったものです。 (Bartley, 1998, Narbonne, 1997, Hagadorn, 1998, McMenamin, 1996)。 現代の刺胞動物の中には単純な巣穴を形成できるものがあり、巣穴痕跡化石の中には二枚貝ではなく刺胞動物が形成したものがある可能性がある(Jensen, 2005)。 いくつかのミミズの巣穴には糞があり、これはより進化したミミズに見られるような消化システムを持っていることを示唆している(Morris, 1993a; Valentine 1994)。 ミミズの胎児の化石は、カンブリア紀初期から発見されています(Bengston, 1997)。
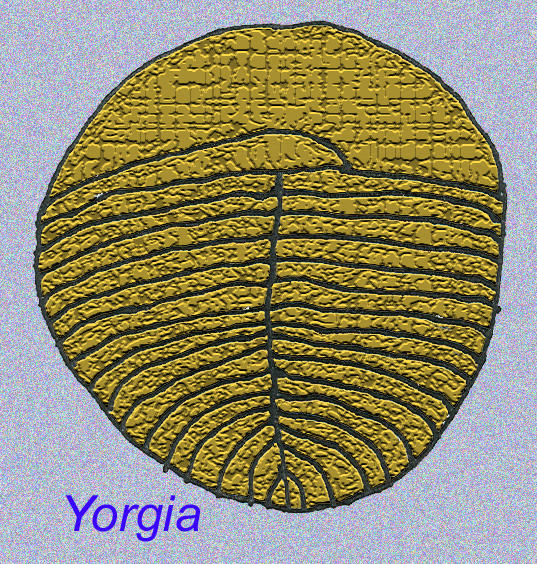
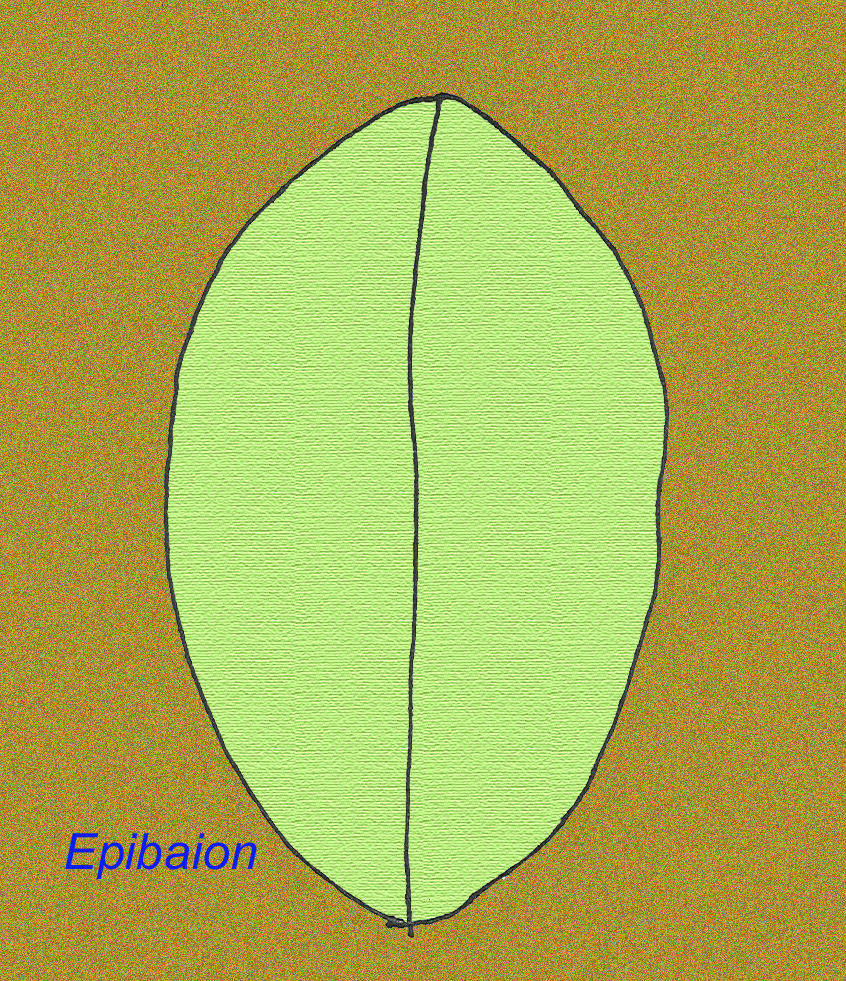
EDIACARAN FAUNAの中には、ワームよりも進化した動物がいるのでしょうか?
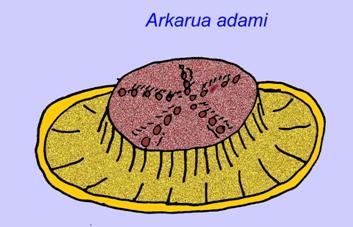
いくつかの遺伝学的な分析では、カンブリア紀以前に腔腸動物の主要グループの分離があったことが示唆されています(Gu, 1998)。 この結論を裏付けるように、多くのエディアカラの化石が腔腸動物のグループに分類されています。 アルクルアは、5分岐の対称性を持ち、カンブリア紀の原始的な棘皮動物(エディオステロイド)との類似性がある。 おそらく5つの構造は中央の溝につながる摂食腕であろう(Gehling, 1987)。 2つの属(RedkiniaとSpriggina)は節足動物の系統のベースに近いかもしれない。 Tribrachidiumは片面に3つの付属物を持ち、オーストラリアやヨーロッパで発見されている(Wright, 1997b; McCall, 2006)。 また、エディアカラの動物の中には、3軸(Tribrachidium, Ventagyrus)、4軸(Conomedusies)、8軸(Eoandromeda)のボディプランを持つものがある。 これらを刺胞動物の絶滅した系統と解釈する人もいます(Xiao, 2009)。
エディアカラの化石Ausiaは、泌尿器系に分類されています(Xiao, 2009).
PhylozoonとKimberella(後述)の2種は、節足動物(環形動物)や軟体動物の親戚である可能性があります。 キンベレーラは両性具有で、腸がある可能性がある(Fedonkin, 1997)。 キンベレーラは、高速で泳ぐ海の捕食者として生きていた軟体動物だったようです。 Cloudinaの貝殻に開けられた穴は、Edicaranの軟体動物の捕食者によるものとされている(McCall, 2006)。
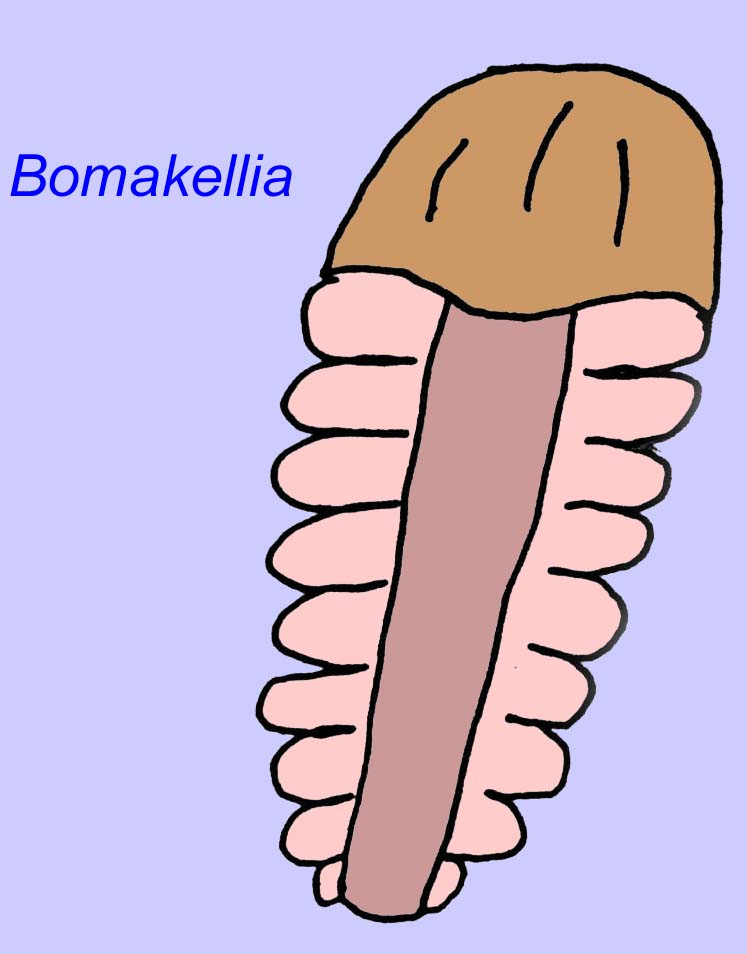
その他の化石は節足動物に関連している可能性があり、そのうちのいくつか(ボマケリアなど)は、三葉虫(古生代の一般的な節足動物の一種;McCall, 2006)のヘッドシールドと比較される構造を持っています。
エディアカラの動物相の中には、3軸(Tribrachidium, Ventagyrus)、4軸(Conomedusies)、8軸(Eoandromeda)のボディプランを持つものがあります。 これらを刺胞動物の絶滅した系統と解釈する人もいます(Xiao, 2009)。

div