De eerste dieren
De meest primitieve levende wezens zijn eencellig: het hele organisme bestaat uit één enkele cel. Sommige organismen kunnen leven als afzonderlijke cellen of als groepen cellen in een kolonie. Als de cellen van een kolonie worden gescheiden, kunnen ze verder leven als afzonderlijke cellen of een nieuwe kolonie vormen. Geen enkele cel is gespecialiseerd voor een specifieke taak. De moderne alg Volvox is hier een goed voorbeeld van: de foto hiernaast is van een Volvox-kolonie.
Multicellulariteit is enkele tientallen keren onafhankelijk van elkaar geëvolueerd in planten, rode algen, bruine algen, schimmels en dieren. Zo omvatten de roodwieren in de Volvocales groep eencellige soorten (Chlamydomonas), gegeneraliseerde viercellige soorten (Basichlamys), en soorten die kolonies vormen die wel 50.000 cellen kunnen omvatten (Volvox) (Sachs, 2008).
Terwijl veel algen in deze eenvoudige kolonies leven, hebben sommige algen wel degelijk cellen die zich specialiseren om verschillende taken uit te voeren (zij hebben weefsels die op wortels lijken en op bladeren).
De eerste fossielen van algen zijn bekend van ongeveer 2,1 miljard jaar geleden en tegen ongeveer 1 miljard jaar geleden is er een toename in de diversiteit van de bekende algensoorten. Eukaryote algen lijken zich plotseling te hebben gediversifieerd rond 600 miljoen jaar geleden, na de ijstijden aan het eind van het Proterozoïcum (Cavalier-Smith, 2000).
Tegen 570 miljoen jaar geleden zijn er algen bekend met een verscheidenheid aan anatomische en reproductieve kenmerken (Xiao, 1998). In het fossielenbestand is een verscheidenheid aan prehistorische algen bewaard gebleven, zoals die welke hieronder zijn afgebeeld.
FUNGI
Veel van de pre-Ediacaran fossielen vertegenwoordigen eenvoudige meercellige kolonies van bacteriën, eukaryote algen (waaronder groene en rode algen), en vormen die gelijkenissen vertonen met schimmels. (Butterfield, 2009; Xiao, 2009).
Carnivore schimmels die hyphale ringen vormen om op nematoden te jagen, zijn waargenomen in 100 miljoen jaar oud barnsteen (Schmidt, 2007).
De EERSTE DIEREN
De Ediacaran fauna is een verzameling organismen genoemd naar de Ediacara heuvels in Australië waar ze voor het eerst werden gevonden. Ediacaran fossielen zijn bekend uit meer dan 30 landen en alle continenten behalve Antarctica en Zuid-Amerika (De, 2005; McCall, 2006).Enkele duizenden specimens, bestaande uit meer dan 270 soorten, zijn beschreven van de Ediacaran fauna en vele moeten nog beschreven worden (Fedonkin, 1990; Minnerop, 2000; Waggoner, 1998; Shen, 2008). Het eerste Ediacaraanse dier werd beschreven in 1872, gevolgd door een tweede vondst in 1908-14 (McCall, 2006).
- ZIJ ZIJN OUDE
Fossiele lagen die zijn gedateerd op meer dan 1,2 miljard jaar oud bevatten dieren – zij bevatten niet alleen fossielen van radiaal-zachte organismen, zij bevatten holen die lijken te zijn gemaakt door slijm-producerende wormen (Rasmussen, 2002; McCall, 2006). Er is een vindplaats in India waarvan de wormholen op 1,1 miljard jaar geleden zijn gedateerd, hoewel sommigen deze datum betwisten. Onlangs is een andere vindplaats met duidelijke tekenen van primitieve dieren (verwanten van kwallen) en wormholen op zijn jongst gedateerd op 1,2 miljard jaar oud (Seilacher, 1998; Kerr, 1998b; Morris, 1993). Andere Ediacaran-fossielen zijn bekend van 610-600 miljoen jaar geleden en meer diverse assemblages van organismen zijn bekend van 555,3 miljoen jaar geleden (McMenamin, 1996, Minnerop, 2000).Precambrische gesteentelagen van 580 miljoen jaar oud bezitten fossielen van kleine bilateranen; grotere bilateran-dieren zijn aanwezig na dateringen van 555 miljoen jaar geleden (Condon, 2005).Zij werden grotendeels vervangen door de dieren van de “Cambrische explosie” die ongeveer 545 miljoen jaar geleden begon. De Ediacaran Periode wordt gedefinieerd als 635 miljoen jaar geleden na de Marinoan Sneeuwbal aarde ijstijd (hoewel de Gaskiers ijstijd van 562 miljoen jaar geleden plaatsvond binnen de Ediacaran Periode) (Xiao, 2009).
Moleculaire vergelijkingen van moderne organismen ondersteunen ook de conclusie dat dieren vóór het Cambrium een verscheidenheid aan lijnen hadden voortgebracht. Eén studie van 129 eiwitten suggereert bijvoorbeeld dat eukaryoten ontstonden tussen 950 en 1.260 miljoen jaar geleden, dieren ontstonden tussen 761 en 957 miljoen jaar geleden, en coelomate lineages scheidden zich tussen 642 en 761 miljoen jaar geleden (Douzery, 2004). Andere analyses hebben soortgelijke resultaten opgeleverd (Peterson, 2004). Andere moleculaire schattingen voor de oorsprong van eumetazoan dieren (cnidarians en bilateans), afkomstig van moleculaire klokstudies, variëren van 570 miljoen jaar geleden tot 1.100 miljoen jaar geleden. De oorsprong van de bilateraantjes is geschat op 580 miljoen jaar geleden (Peterson, 2005). Fossielen van dieren gevonden in Australië, India en Texas zijn gedateerd op meer dan 1 miljard jaar oud en een gesegmenteerd dier uit Rusland is gedateerd van 890 miljoen tot 1 miljard jaar oud (De, 2005). Vanwege de spannende ontdekkingen van dieren uit het Precambrium werd een nieuwe periode toegevoegd aan de geologische tijdschaal, wat de eerste keer is dat een nieuwe periode stratigrafisch is gedefinieerd sinds 1891 (toen het Carboon werd opgedeeld). De Ediacaran periode is gedefinieerd als de tijd tussen 630 miljoen jaar geleden en het begin van het Cambrium, ongeveer 542 miljoen jaar geleden (Knoll, 2004).
Hoe belangrijk zijn deze data? Tot voor kort dacht men dat een verscheidenheid aan complexe dieren plotseling verscheen in de “Cambrische explosie” die ongeveer 545 miljoen jaar geleden begon. We weten nu dat dieren al minstens 60 miljoen jaar daarvoor bestonden en misschien wel 600 miljoen jaar voor het Cambrium. Zestig miljoen jaar lijkt misschien niet zo lang gezien de uitgestrekte tijdspanne in het Precambrium, maar in zo’n periode kan heel wat evolutie van dieren plaatsvinden. Het duurde ongeveer zestig miljoen jaar voor de rhipidistische vissen evolueerden tot de eerste amfibieën en ongeveer zestig miljoen jaar voor de daaropvolgende diversificatie van deze amfibieën in een verscheidenheid van lijnen waaronder de eerste reptielen. Een periode van zestig miljoen jaar, beginnend aan het eind van het Perm, zou twee massa-uitstervingen omvatten. Het begint met een wereld die gedomineerd wordt door synapside reptielen en primitieve archosauriërs en eindigt in het Jura met zijn diverse lijnen van dinosauriërs, pterosauriërs en mariene reptielen. Over ongeveer zestig miljoen jaar zijn de primitieve zoogdieren die het uitsterven van de dinosauriërs overleefden, uitgegroeid tot de diverse fossiele en levende groepen van het Cenozoïcum.


2) ZE ZIJN EENVOUDIGE DIEREN
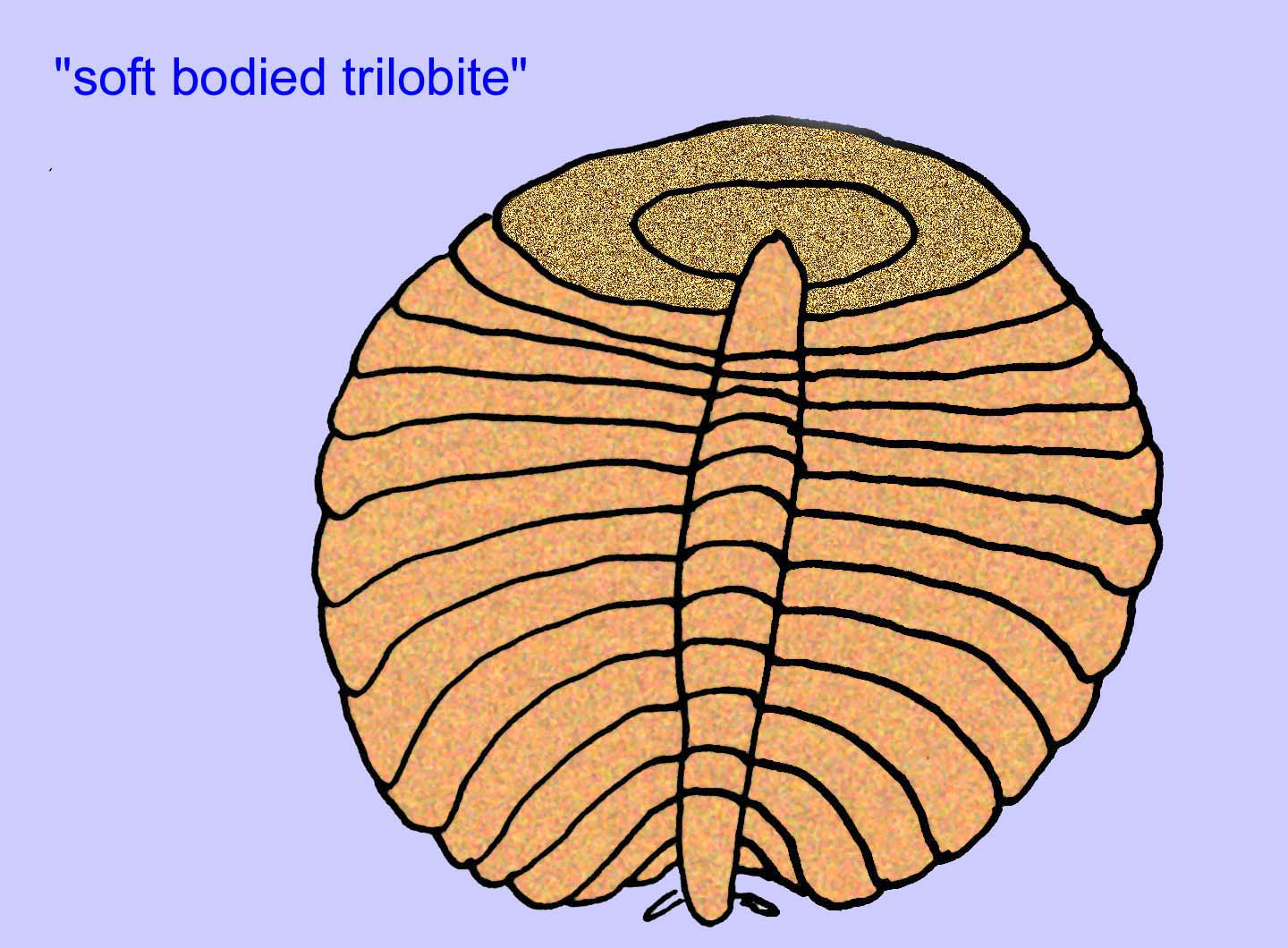
De meeste Ediacaran-organismen zijn minder dan een centimeter groot, maar sommige zijn wel een paar meter lang. Ze zijn vreemd en de meeste verschillen sterk van alles wat tegenwoordig leeft (velen noemen ze de groep Vendobionta). Vrijwel alle hebben geen hoofd of staart, binnen- of buitenkant, voor- of achterkant, tanden of ogen. De meeste hebben geen duidelijke bloedsomloop, zenuwstelsel, spijsverteringsstelsel of inwendige organen van welke soort dan ook. De meeste zijn zeer dun en plat – een toestand die essentieel zou zijn voor dieren zonder bloedsomloop, ademhalingsstelsel, spijsverteringsstelsel of uitscheidingsstelsel. Sommige vindplaatsen hebben een grote verscheidenheid aan dieren, met wel 30 verschillende taxa (Buss, 1994).
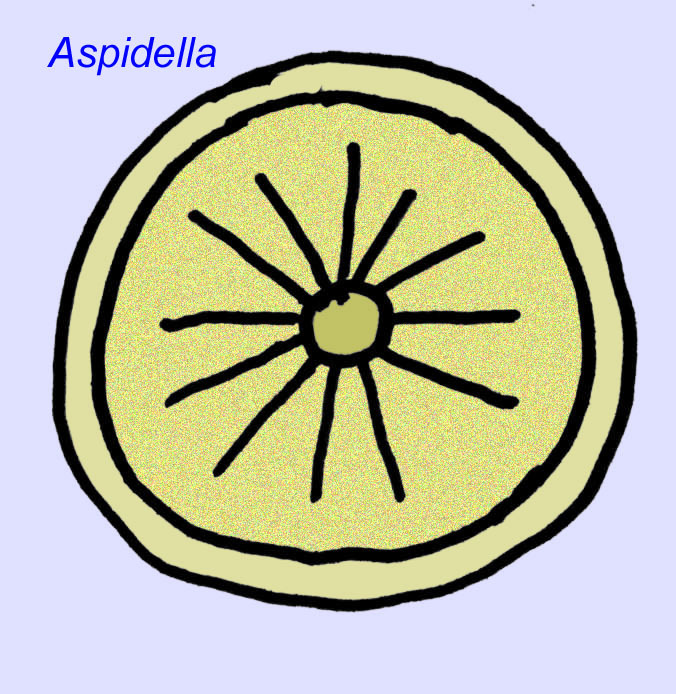
Wat zijn dat? Er zijn in het verleden verschillende meningen geweest over deze organismen: sommigen classificeerden ze als planten (de conclusie vanaf de Eerste Wereldoorlog), zeer grote eencellige organismen, unieke korstmossen, een geheel uitgestorven groep dieren, en een groep dieren die de voorouders van de moderne dieren omvat. Oorspronkelijk dacht men dat ze uitgestorven waren lang voor de opkomst van moderne diergroepen, nu is het duidelijk dat sommige groepen overleefden tot in het Cambrium.Sommige kunnen fossielen zijn van niet-metazoan dieren (Droser, 2006). De dichtheid van individuen van het schijfvormige dier Aspidella wijst erop dat de biomassa van bodemdieren in het Precambrium die kan hebben benaderd die in moderne gemeenschappen wordt waargenomen (Gehling, 2000).
Veel van de pre-Ediacarische fossielen zijn eenvoudige meercellige kolonies van bacteriën, eukaryote algen (waaronder groene en rode algen), en vormen die gelijkenis vertonen met schimmels. Sommige fossielen uit het Neoproterozoïcum zijn geïnterpreteerd als de spicules en larven van sponzen (Butterfield, 2009).
Sponzen, die tot de eenvoudigste dieren behoren, zijn geïdentificeerd in Ediacaraanse sedimenten (zoals het geslacht Palaeophragmodictya) die gelijken op sommige Paleozoïsche sponzen. Deze sponzen konden 1,5-10 cm in diameter meten (Gehling, 1996; Kerr, 1998a). Sponsspicules zijn aanwezig in Ediacaran gesteenten (Xiao, 2000).
Sommige Ediacaran fossielen zijn geïnterpreteerd als schimmels en basale dieren die primitiever zijn dan sponzen (Xiao, 2009).
CNIDARIAAN
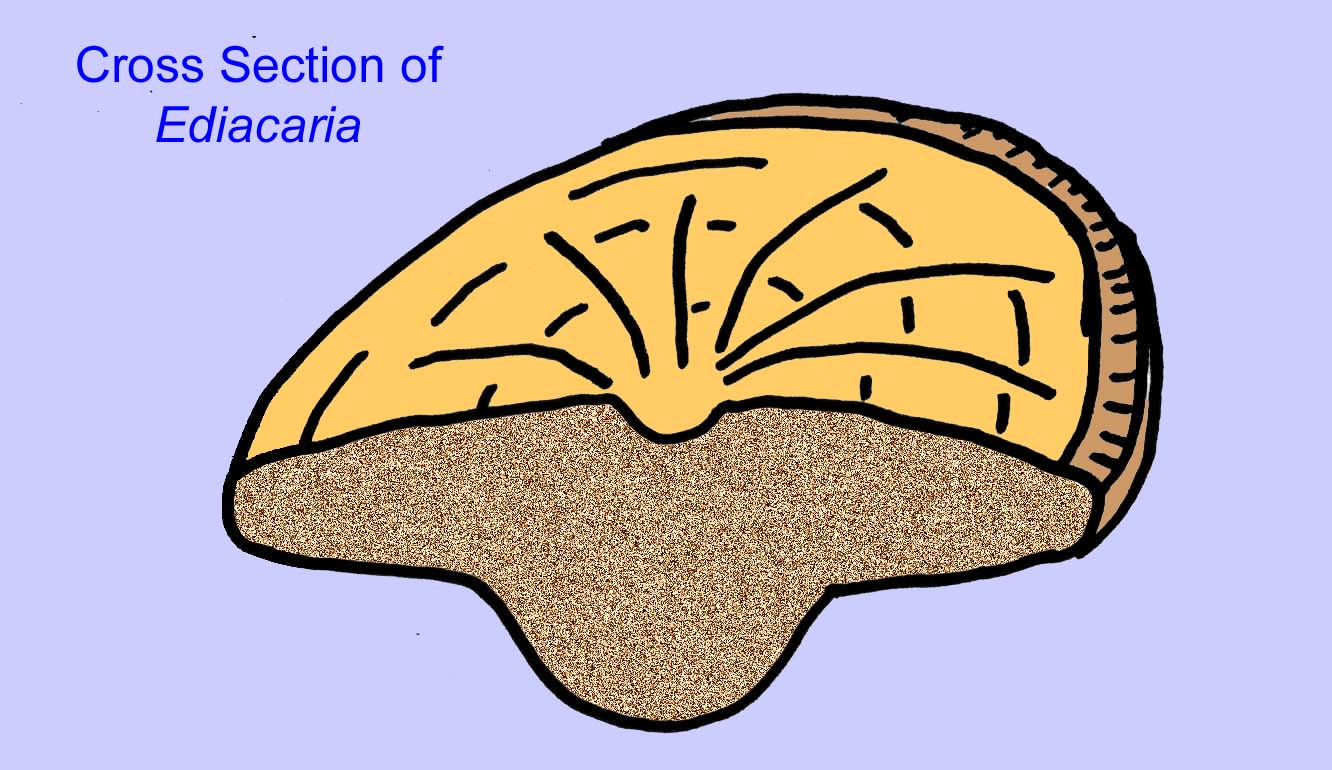
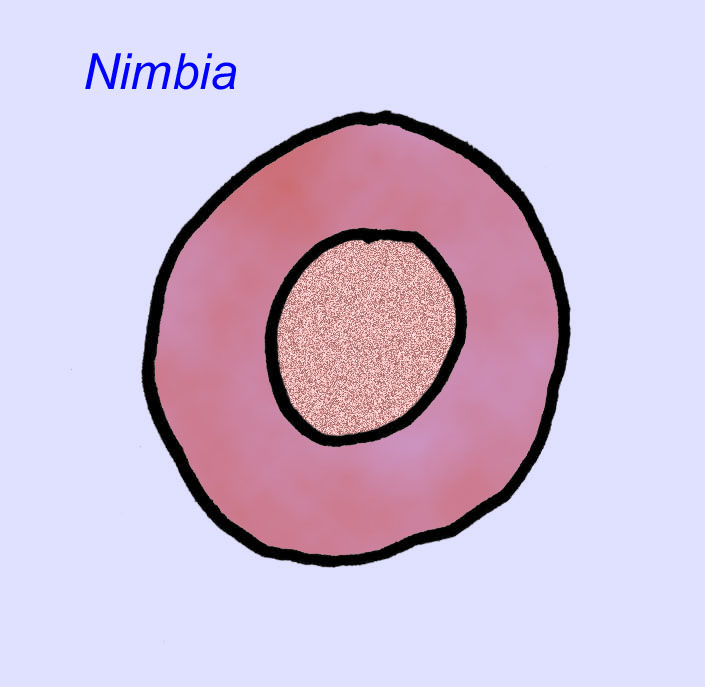
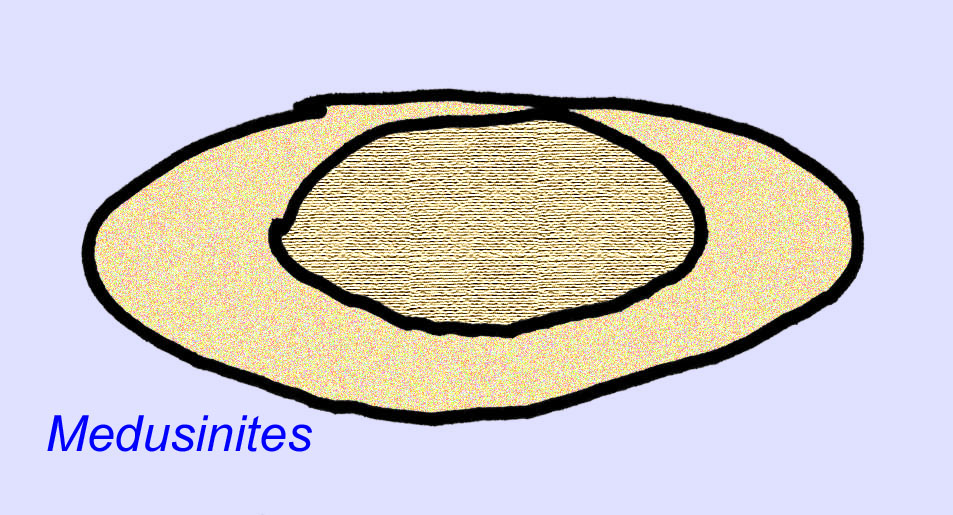
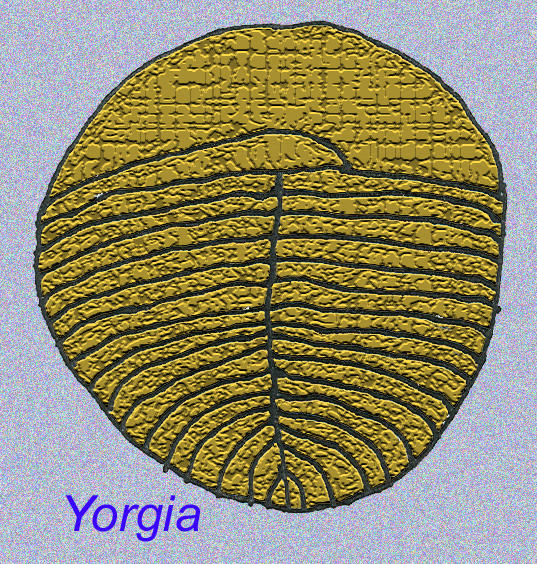
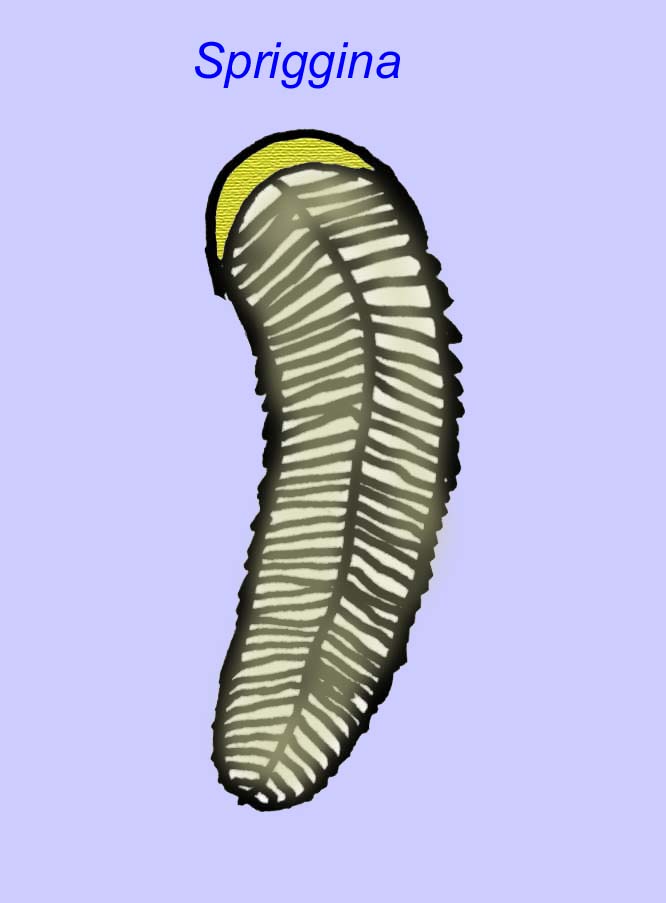
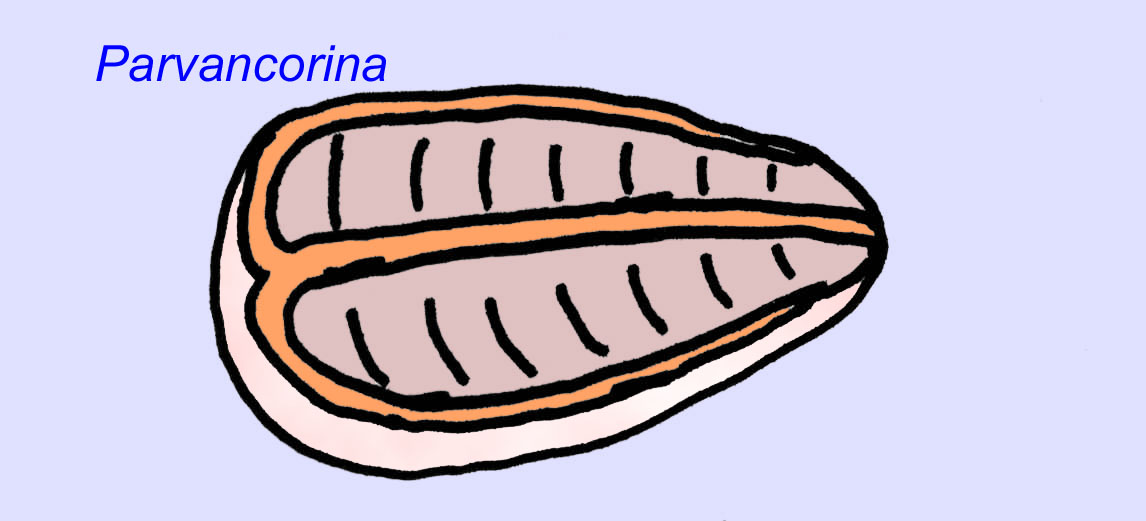
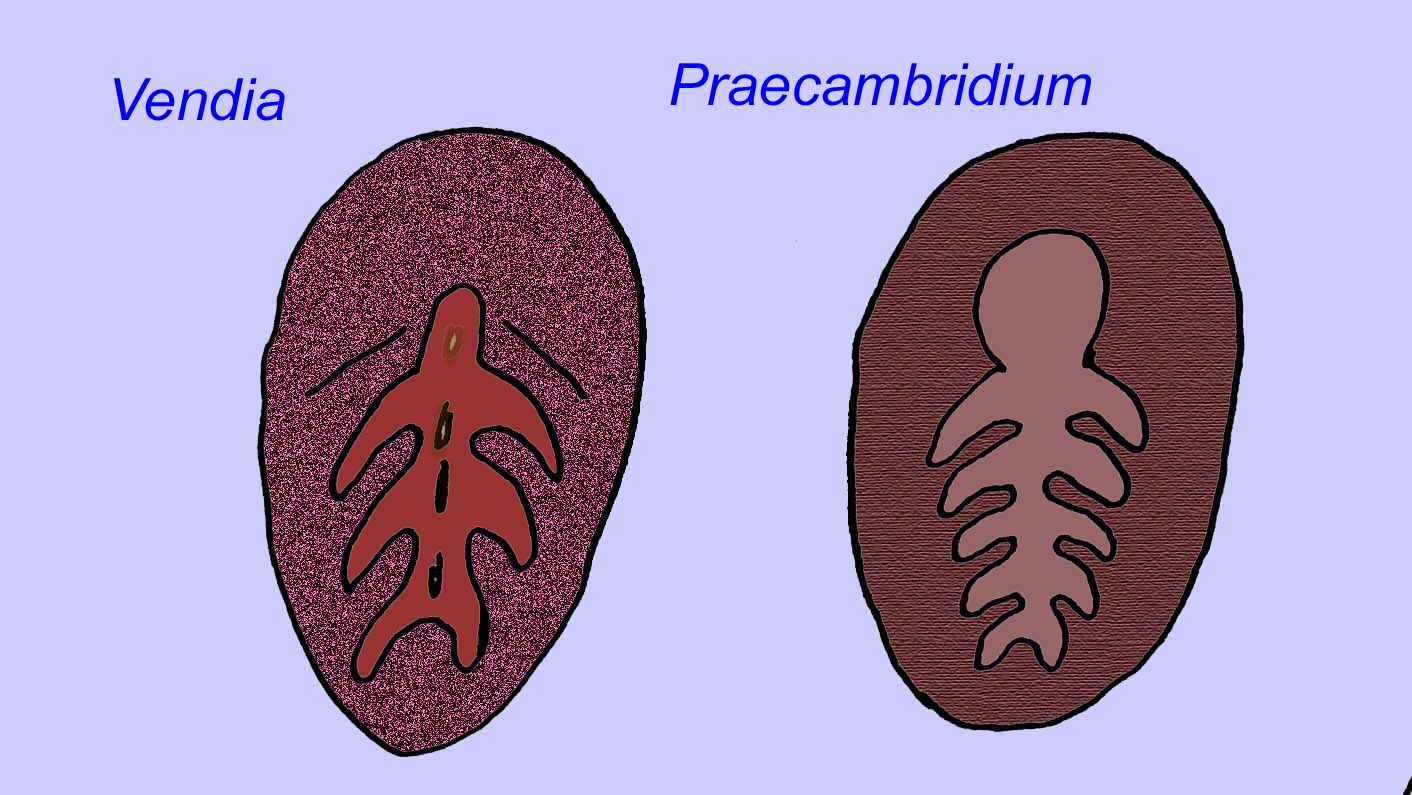
Er is een algemene consensus dat veel Ediacaran-fossielen moeten worden geclassificeerd als vroege cnidarians, een groep dieren waartoe de moderne kwallen, zeenaalden en zeewaaiers behoren. Sommige vormen (zoals Ediacaria en Nimbia) zijn radiaal symmetrisch en lijken oppervlakkig gezien op cnidariërs, maar missen elk teken van een mond, hetgeen een reden kan zijn om ze van deze groep uit te sluiten. (Het is niet zeker of de eerste cnidariërs een mond zouden hebben gehad, zoals alle latere vormen hebben). Andere, zoals Albumares brunsae, bezitten een gastrovasculair systeem dat lijkt op dat van cnidarianen (Lipps, 1992; Sokolov, 1985). Vele hebben een centrale schijfvormige structuur aan één kant van hun platte, ronde lichaam (Crimes, 1995; Bertrand-Sarfati, 1995). Sommige Ediacaran-dieren (Spriggina, Parvancorina, en Praecambridium) hebben een onderscheid tussen hun boven- en onderzijde (achter- en voorzijde). Sommige, zoals Ediacaria, hadden radiale lijnen en perifere verdikkingen (Crimes, 1995). Sommige van de Ediacaraanse cnidaria zijn geclassificeerd als een aparte maar uitgestorven subklasse (McCall, 2006).
Wel ontbrak het deze dieren aan een gemineraliseerd skelet, bezaten de meeste of zelfs alle dieren (zoals Cyclomedusa en Ernietta) een soort stijf skelet dat in staat was een afdruk in de modder te maken. Het type weefsel waaruit het inwendige skelet bestond is niet bekend (Crimes, 1995; Dzik, 1999).
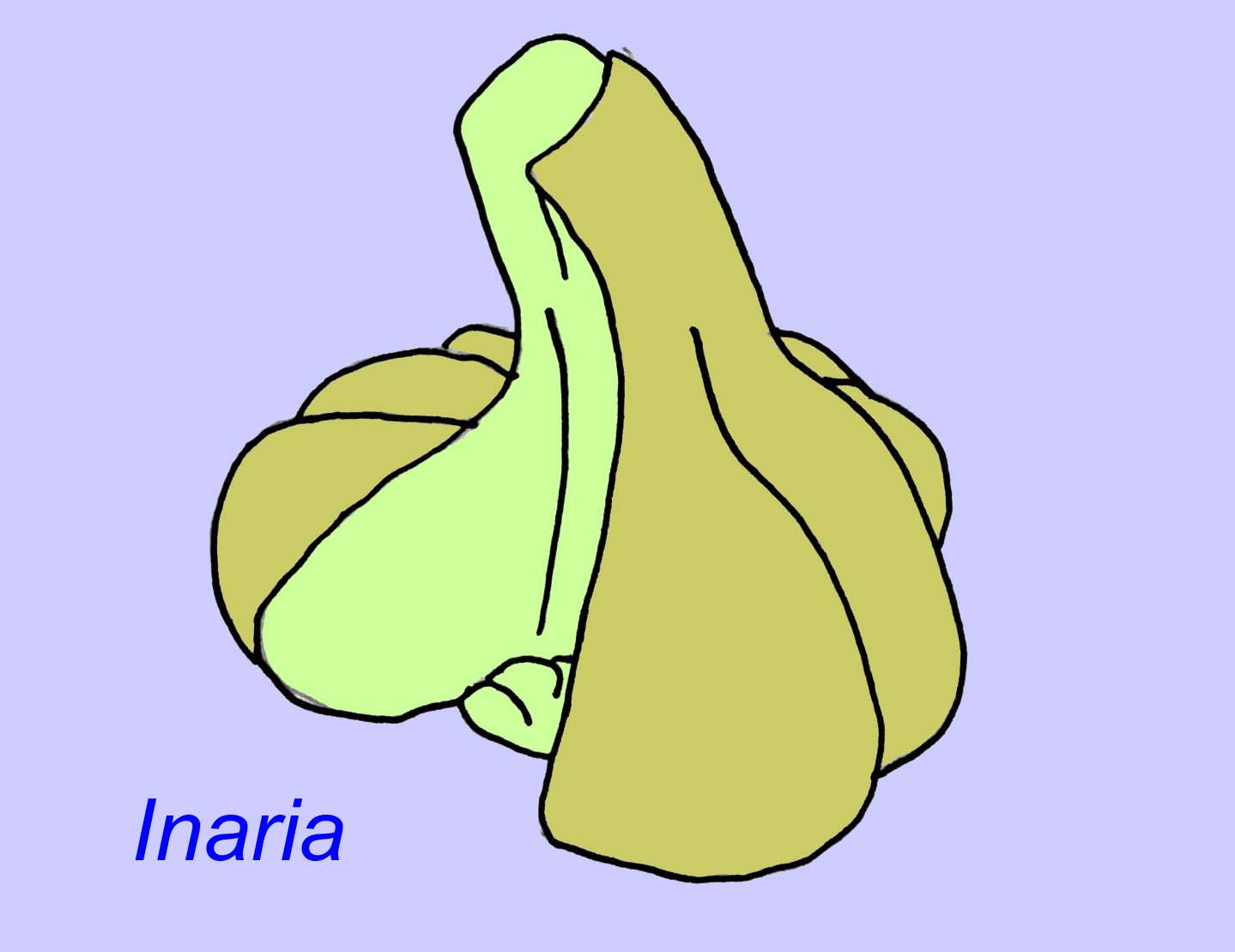
Mackenzia lijkt ook een cnidarian te zijn (zelfs als dat niet zo is, is het vrijwel zeker een metazoïsch dier). Er zijn indirecte aanwijzingen voor voedselbolussen in een spijsverteringsholte en voor musculatuur (Morris, 1993a). Sinocyclocyclicus kan een basale cnidarian (of zelfs een basale metazoan) zijn waarvan de apicale knopvorming vergelijkbaar is met die van sommige cnidarians (zoals tabulate koralen) (Xiao, 2000). Inaria was een cnidariër van de klasse Anthozoa met een mondrand, een keelholte, en een kamer waarin zich fotosynthetiserende organismen kunnen hebben bevonden zoals die in veel moderne koralen worden aangetroffen (McMenaman, 1990, Gehling, 1988).
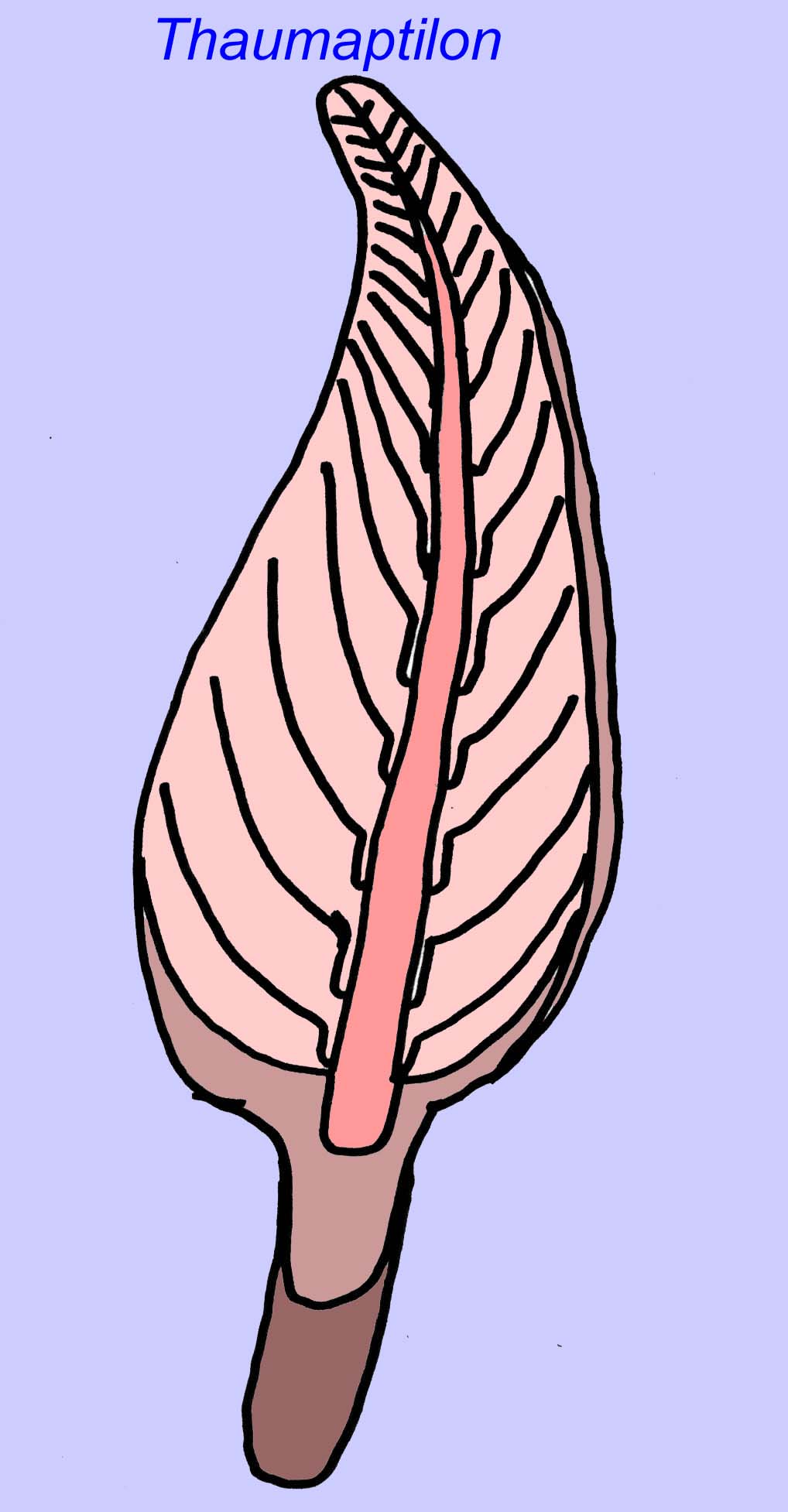
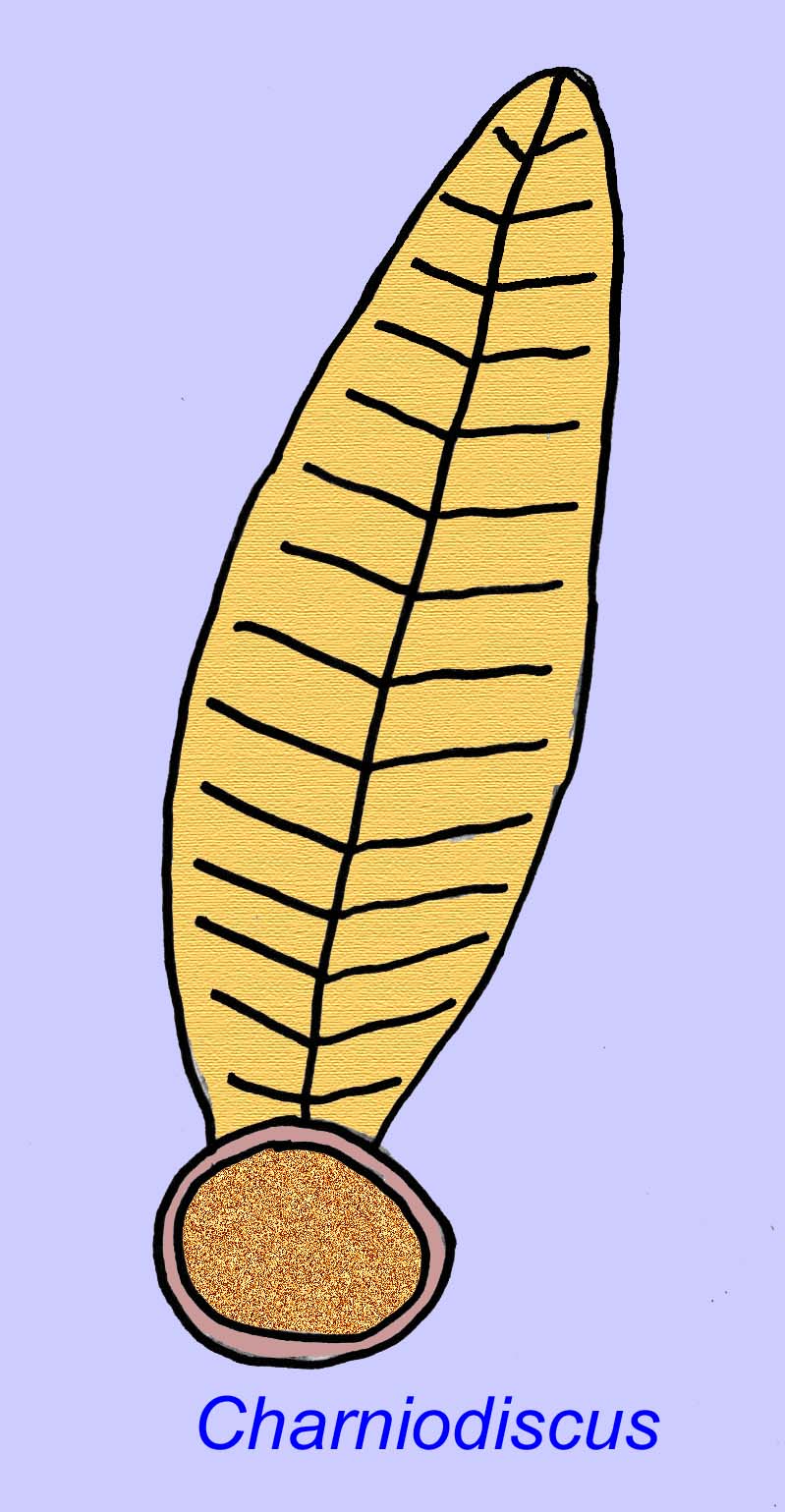
Thaumaptilon was een voorvorkachtig dier dat verwant lijkt aan de moderne zeeduivels. Het voedde zich met puin en gebruikte een houvast om zich op de bodem vast te zetten (Morris, 1993a; 1993b). De frond van Charniodiscus kon een lengte bereiken van 1,2 meter (Sokolov, 1990).
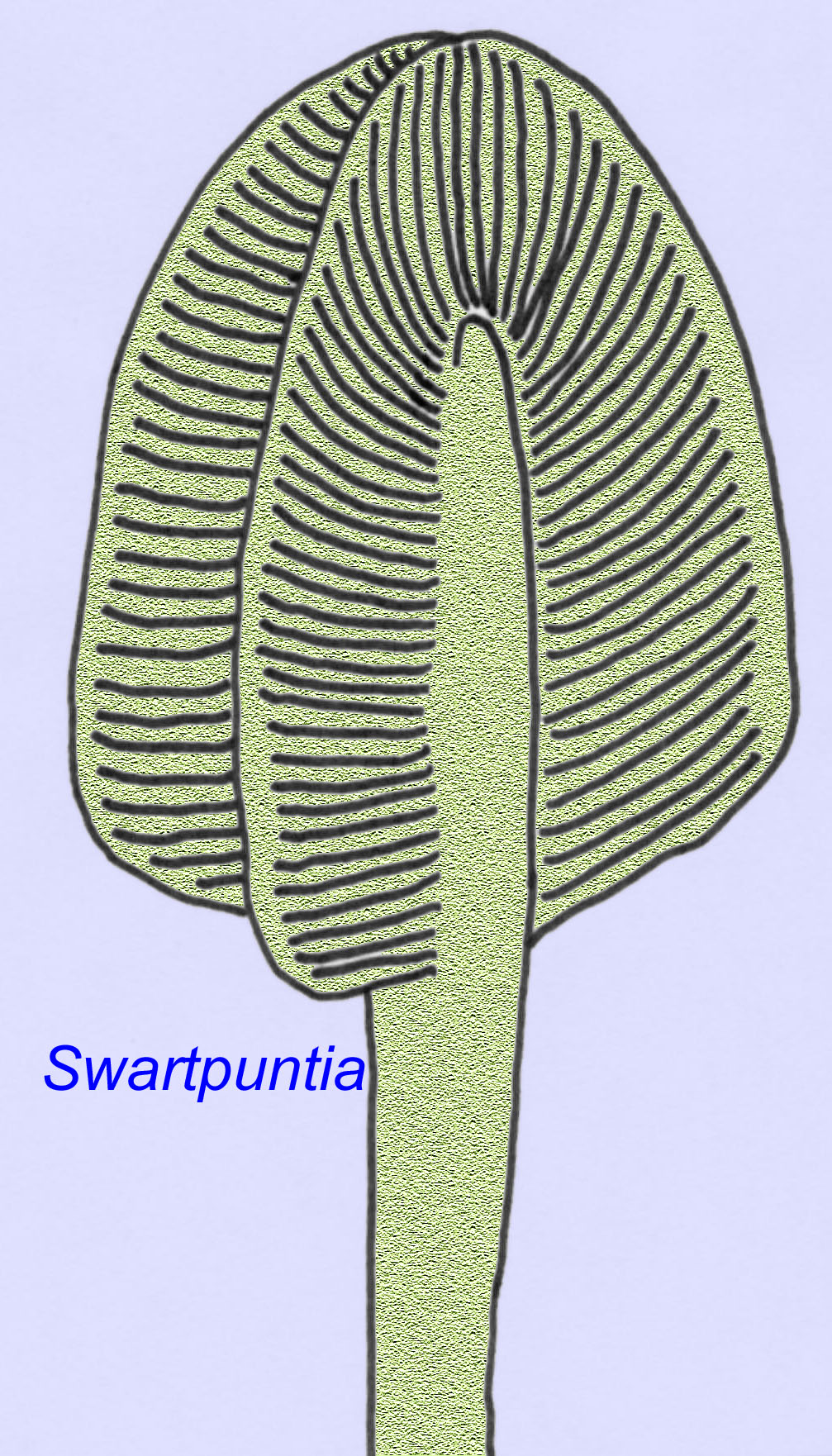
Vele Ediacaran-dieren (zoals Pteridinium, Swartpuntia, Charnia, en Charniodiscus) bezitten een rechtopstaande stengel die verbonden is met gesegmenteerde petaloïden.
Een exemplaar van Charnodiscus mat 1,2 meter in lengte.
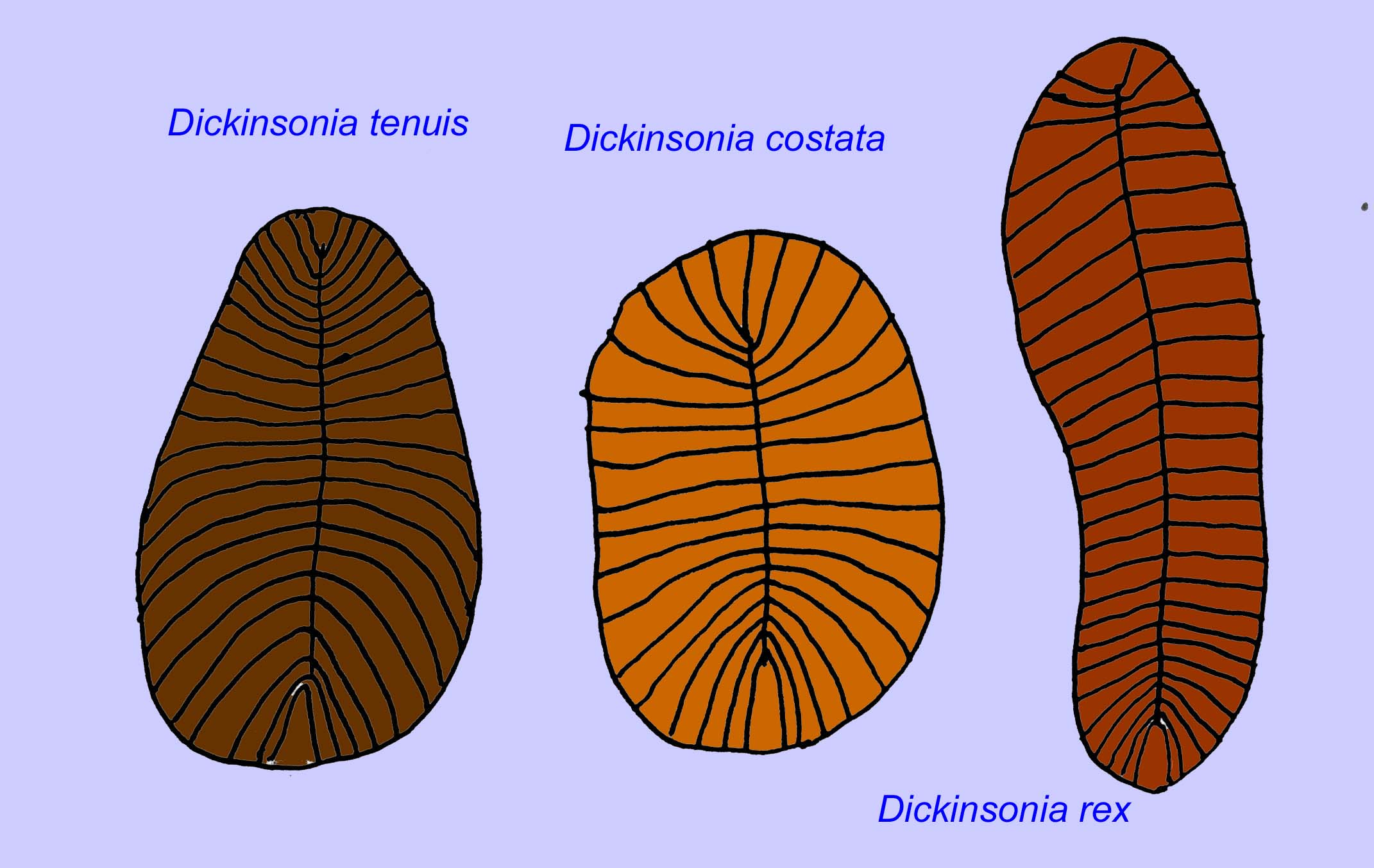
De microscopische anatomie van Swartpuntia lijkt sterk op die van Dickinsonia, een vroege worm (Narbonne, 1997; McCall, 2006). Gefossiliseerde embryo’s die lijken te behoren tot bilateran dieren zijn bekend van 570 miljoen jaar geleden (Xiao, 1998).

Wormen
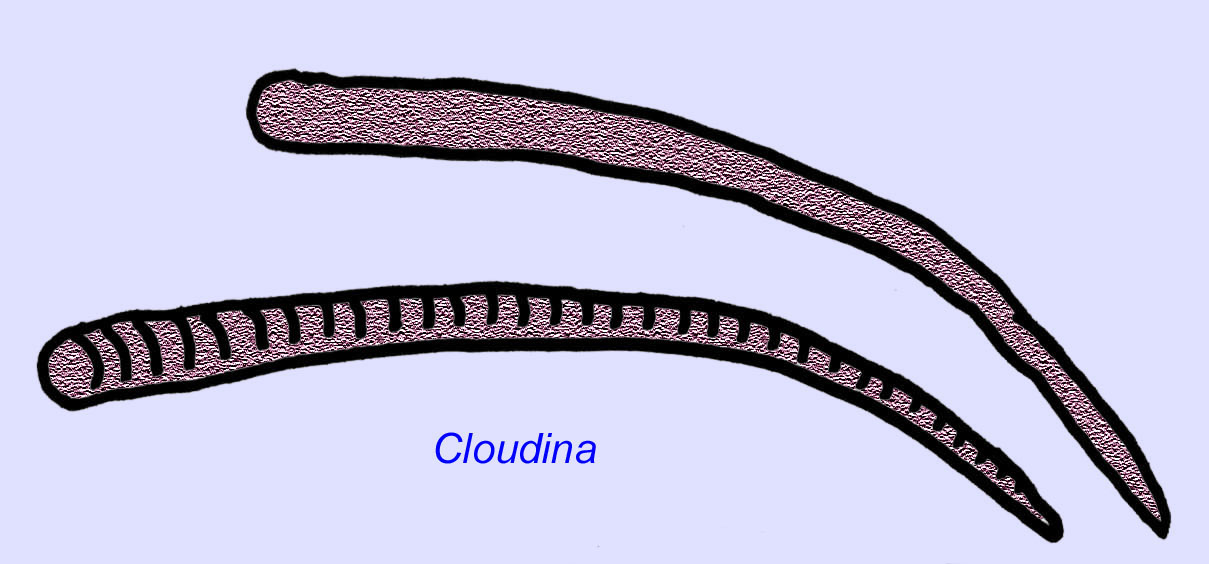
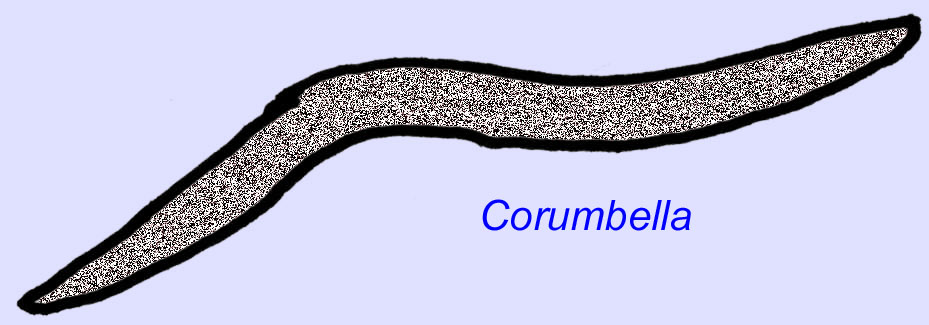
Ediacaran wormen zijn bekend uit Azië, Europa, Afrika, Australië en Noord-Amerika (McCall, 2006). Een aantal Ediacaran-dieren was buisvormig, zoals Archaeichnium, Cloudina, Corumbella, en Onuphionella. Terwijl Onuphionella is vergeleken met annelid wormen, werd Corumbella vergeleken met de buizen gebouwd door de poliepen van sommige cnidarianen. Terwijl Archaeichnium flexibel was, was Cloudina bedekt met een gemineraliseerd skelet. Sommige buizen van Cloudina (maximumlengte minder dan 4 cm) waren geannuleerd en andere waren glad, hoewel dit kan wijzen op verschillen in conservering (Hagadorn, 2000). Cloudina is het eerste bekende dier met een gemineraliseerd skelet en in 3% van de specimens zijn gaten geboord die wijzen op predatie. Het is mogelijk dat deze predatie de hardere structuren, die zo kenmerkend zijn voor veel van de dieren uit het Cambrium, heeft bevorderd (Bengston, 1992). Cloudina is bekend uit het Precambrium van Zuid-Amerika, Europa, het Midden-Oosten en Zuidoost-Azië en verwanten zijn bekend uit het Vroege Cambrium (vóór de verspreiding van trilobieten) in Noord-Amerika (Grant, 1990). Fossielen van Cloudina zijn bekend van ten minste 6 miljoen jaar voor het begin van het Cambrium (Condon, 2005).
Naast de twee erkende soorten Cloudina bezaten nog een aantal andere fossiele soorten verkalkte schelpen in het Late Precambrium (McCall, 2006). Cloudina, Namacalanthus, en Conotubus zijn geskeletteerde Precambriaanse dieren die ook bekend zijn in het vroege Cambrium (Shu, 2008).
Een aantal wormen zoals Paleolina evenkiana en Planolites beverlyensis zijn aan het eind van het Proterozoïcum en tot in het Cambrium gevonden. De fossielen van Planolites zijn 1-2 mm breed en 5 tot 13 mm lang (Bartley, 1998). Niet gesegmenteerde fossielen zoals Protechirus, Platypholina, en Vladimissa worden niet goed begrepen. Sommige fossielen zijn gesegmenteerd, zoals Dickinsonia, die wordt geclassificeerd als een annelid worm. De kleinste exemplaren van Dickinsonia zijn 4 mm in doorsnee, terwijl de grootste 1 meter lang zijn. Ondanks de lengte van deze wormen, waren ze zeer dun, minder dan 3 mm. Er zijn aanwijzingen dat de hoeveelheid zuurstof in de atmosfeer aan het eind van het Precambrium slechts ongeveer een tiende was van de huidige concentratie en de extreme dunheid (en dus het grote oppervlak) van deze wormen kan een aanpassing zijn geweest voor de zuurstofdiffusie. Deze wormen waren algemeen en wijdverspreid. Zij voedden zich met puin op de oceaanbodem, gezien het sediment dat vaak hun darmen vulde (Runnegar, 1982). Het eenvoudige karakter van Dickinsonia kan erop wijzen dat het eerder een cnidarian dan een bilateran is (Brasier, 2008).
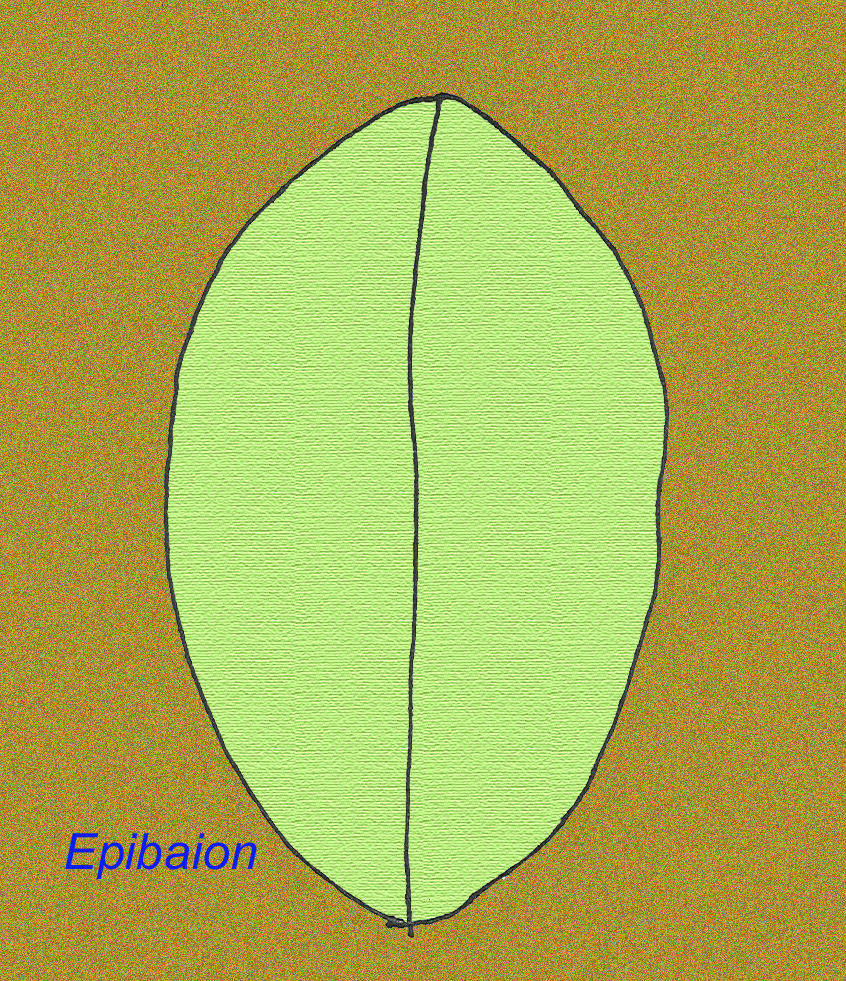
Een groot Ediacran dier, Epibaion axiferus genaamd, heeft indrukken achtergelaten die tot 4,3 meter bij .15 meter reiken (met een tweede exemplaar van 2,5 meter bij .35 meter) (Ivantsov, 2002).


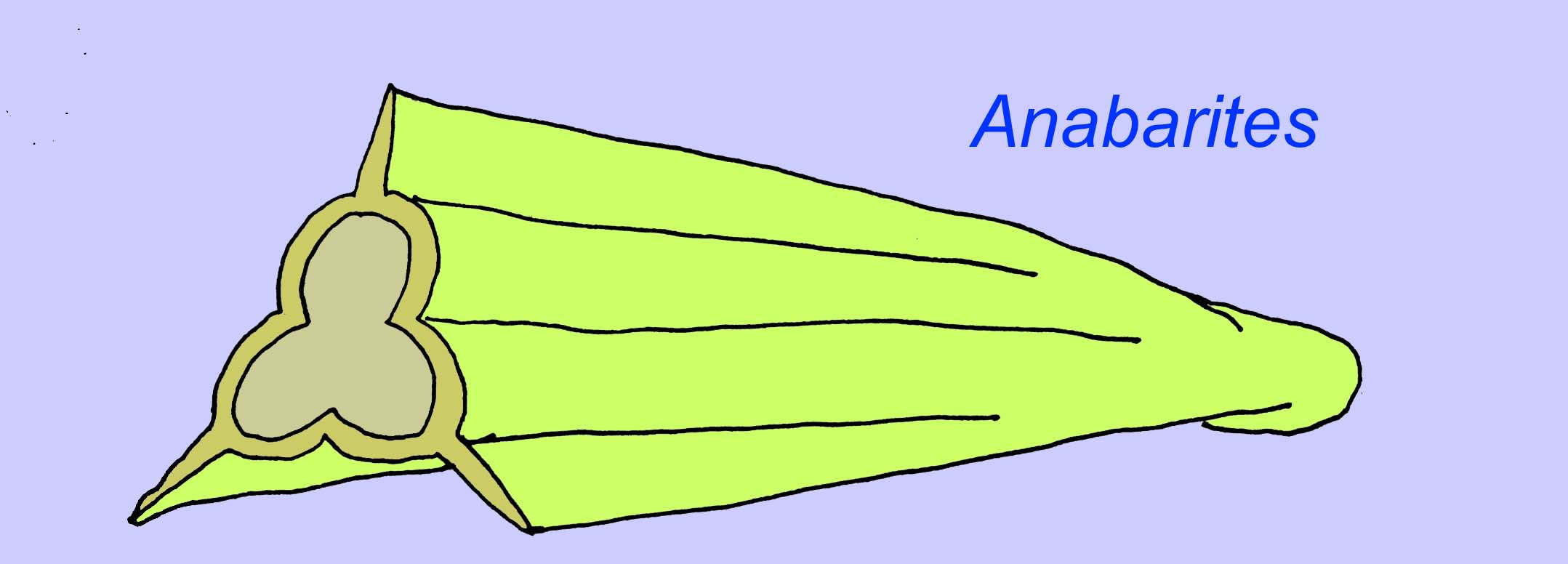
Anabaritiden zijn fossielen met schalen uit het vroege Cambrium. Sommige, zoals Anabarites, dateren mogelijk uit het eind van het Precambrium (McMenamin, 1990).
Op een aantal Proterozoïsche vindplaatsen zijn sporenfossielen van wormholen gevonden. Deze holen werden gemaakt door wormen die verder ontwikkeld waren dan platwormen. (Bartley, 1998, Narbonne, 1997, Hagadorn, 1998, McMenamin, 1996). Sommige moderne cnidariërs kunnen eenvoudige holen vormen en het is mogelijk dat sommige van de holspoorfossielen door cnidariërs in plaats van bilateranen werden gevormd (Jensen, 2005). Sommige wormholen zijn compleet met uitwerpselen, hetgeen wijst op een spijsverteringssysteem zoals dat bij meer gevorderde wormen wordt aangetroffen (Morris, 1993a; Valentine 1994). Fossielen van wormembryo’s zijn gevonden vanaf het vroegste Cambrium (Bengston, 1997).
ZIJN ER BESTAANDE DIEREN DIE MEER GEVORDERD ZIJN DAN WORMEN ONDER DE EDIACARAN FAUNA?
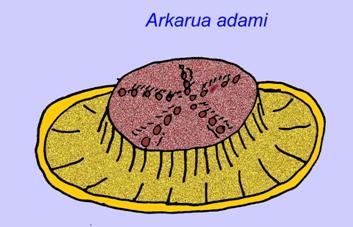
Sommige genetische analyses hebben gesuggereerd dat de scheiding van de belangrijkste groepen coelomaatdieren vóór het Cambrium plaatsvond (Gu, 1998). Een aantal Ediacaran-fossielen zijn bij coelomaatgroepen ingedeeld, hetgeen deze conclusie ondersteunt. Arkurua vertoont overeenkomsten met de stekelhuidigen met zijn 5-vertakte symmetrie en zijn gelijkenis met primitieve stekelhuidigen uit het Cambrium (de edrioasteroïden). Misschien zijn de 5 structuren voedingsarmen die naar een centrale groef leiden (Gehling, 1987). Twee geslachten (Redkinia en Spriggina) staan wellicht dicht bij de basis van de arthropoden-lijnen. Tribrachidium, met drie aanhangsels op één van zijn oppervlakken, is gevonden in Australië en Europa (Wright, 1997b; McCall, 2006). Een deel van de Ediacaran fauna had een lichaamsplan dat triradiaal (Tribrachidium, Ventagyrus), tetraradiaal (Conomedusies), en octaradiaal (Eoandromeda) was. Sommigen hebben deze geïnterpreteerd als uitgestorven lijnen van cnidarians (Xiao, 2009).
Het Ediacaran-fossiel Ausia is geclassificeerd als een urochordaat (Xiao, 2009).
Twee soorten, Phylozoon en Kimberella (zie hieronder), zijn mogelijk verwant aan gesegmenteerde wormen (anneliden) of mollusken. Kimberella heeft bilaterale symmetrie en een mogelijke darm (Fedonkin, 1997). Kimberella lijkt een weekdier te zijn geweest dat leefde als een snel zwemmend marien roofdier. De gaten die in de schelpen van Cloudina zijn geboord, zijn toegeschreven aan Edicaraanse weekdierroofdieren (McCall, 2006). Fossielen van embryo’s uit het late Precambrium vormden poollichamen die lijken op veel weekdieren (Chen, 2006).
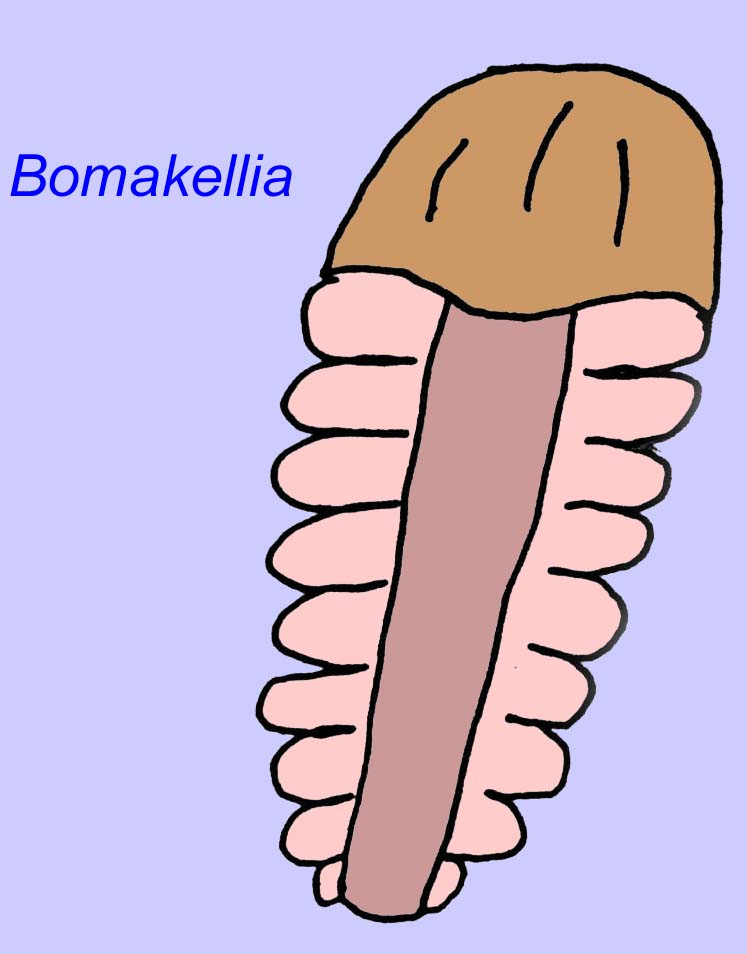
Andere fossielen kunnen verwant zijn aan geleedpotigen, waarvan sommige (zoals Bomakellia) een structuur hebben die is vergeleken met het kopschild van trilobieten (een veelvoorkomend type geleedpotige uit het Paleozoïcum; McCall, 2006).
Sommige Ediacaraanse dieren hadden een driedelig (Tribrachidium, Ventagyrus), tetradiaal (Conomedusies), en octaradiaal (Eoandromeda) lichaamsplan. Sommigen hebben deze geïnterpreteerd als uitgestorven lijnen van cnidarians (Xiao, 2009).