Os Primeiros Animais
Os seres vivos mais primitivos são unicelulares: o organismo inteiro é composto por uma única célula. Alguns organismos podem viver como células individuais ou como grupos de células de uma colónia. Se as células de uma colónia forem separadas, podem continuar a viver como células individuais ou a formar uma nova colónia. Nenhuma célula é especializada para qualquer tarefa específica. A moderna alga Volvox é um bom exemplo disto: a fotografia adjacente é de uma colónia Volvox.
Multicelularidade evoluiu independentemente algumas dezenas de vezes em plantas, algas vermelhas, algas castanhas, fungos, e animais. Por exemplo, as algas re-celulares no grupo Volvocales incluem espécies monocelulares (Chlamydomonas), espécies generalizadas de quatro células (Basichlamys), e espécies que formam colónias que podem incluir 50.000 colónias de células (Volvox) (Sachs, 2008).
Enquanto muitas algas vivem nestas simples colónias, algumas algas têm células especializadas para realizar diferentes tarefas (têm tecidos que são semelhantes às raízes em função e semelhantes às folhas em função).
Os primeiros fósseis de algas são conhecidos de há cerca de 2,1 mil milhões de anos e, há cerca de mil milhões de anos, existe um aumento na diversidade de espécies de algas conhecidas. As algas eucarióticas parecem ter-se diversificado subitamente há cerca de 600 milhões de anos, após o fim da Idade do Gelo Proterozóico (Cavalier-Smith, 2000).
Por 570 milhões de anos atrás, as algas são conhecidas com uma variedade de características anatómicas e reprodutivas (Xiao, 1998). O registo fóssil preservou uma diversidade de algas pré-históricas como as que se descrevem abaixo.
 p>FUNGI
p>FUNGI
MuNGI
Muitos dos fósseis pré-Ediacaran representam simples colónias multicelulares de bactérias, algas eucarióticas (incluindo algas verdes e vermelhas), e formas que têm semelhanças com fungos. (Butterfield, 2009; Xiao, 2009).
Fungos carnívoros formando anéis hifais para presas em nemátodos foram observados em âmbar com 100 milhões de anos (Schmidt, 2007).
Os PRIMEIROS ANIMAIS
A fauna Ediacaran é um conjunto de organismos com o nome das colinas de Ediacara na Austrália onde foram encontrados pela primeira vez. Os fósseis de Ediacaran são conhecidos de mais de 30 países e de todos os continentes excepto Antárctida e América do Sul (De, 2005; McCall, 2006).Vários milhares de espécimes, compreendendo mais de 270 espécies, foram descritos da fauna Ediacaran e muitos ainda não foram descritos (Fedonkin, 1990; Minnerop, 2000; Waggoner, 1998; Shen, 2008). O primeiro animal de Ediacaran foi descrito em 1872, seguido por uma segunda descoberta em 1908-14 (McCall, 2006).
- SÃO VELHOSOS
Estratos fósseis datados de há mais de 1,2 mil milhões de anos contêm animais – não contêm apenas fósseis de organismos de corpo macio radial, contêm tocas que parecem ter sido feitas por vermes produtores de muco (Rasmussen, 2002; McCall, 2006). Existe um sítio na Índia cujas tocas de vermes foram datadas há 1,1 mil milhões de anos, embora alguns contestem esta data. Recentemente, outro sítio com sinais aparentes de animais primitivos (parentes de medusas) e tocas de vermes foi datado com 1,2 mil milhões de anos de idade nos mais novos (Seilacher, 1998; Kerr, 1998b; Morris, 1993). Outros fósseis de Ediacaran são conhecidos há 610-600 milhões de anos e conjuntos mais diversos de organismos são conhecidos há 555,3 milhões de anos (McMenamin, 1996, Minnerop, 2000). Os estratos rochosos pré-cambrianos com 580 milhões de anos possuem fósseis de pequenos bilateranos; animais bilateranos maiores estão presentes após datas de 555 milhões de anos (Condon, 2005). O Período Ediacaran é definido como 635 milhões de anos atrás, após a glaciação Marinoan Snowball earth (embora a glaciação Gaskiers de 562 milhões de anos atrás tenha ocorrido no Período Ediacaran) (Xiao, 2009).
Comparações moleculares de organismos modernos também apoiam a conclusão de que os animais tinham produzido uma variedade de linhagens antes do Cambriano. Por exemplo, um estudo de 129 proteínas sugere que os eucariotas surgiram entre 950 e 1.260 milhões de anos atrás, os animais surgiram entre 761 e 957 milhões de anos atrás, e as linhagens coelomatosas separadas entre 642 e 761 milhões de anos atrás (Douzery, 2004). Outras análises produziram resultados semelhantes (Peterson, 2004). Outras estimativas moleculares para a origem dos animais eumetazoários (cnidários e bilateanos) fornecidas por estudos de relógio molecular variaram entre 570 milhões de anos atrás e 1,100 milhões de anos atrás. A origem dos animais bilateranos foi estimada em 580 milhões de anos atrás (Peterson, 2005). Fósseis de animais encontrados na Austrália, Índia e Texas foram datados com mais de 1 bilião de anos e um animal segmentado da Rússia foi datado com 890 milhões a 1 bilião de anos (De, 2005). Devido às excitantes descobertas dos animais pré-cambrianos, foi acrescentado um novo período à escala temporal geológica, que é a primeira vez que um novo período foi definido estratigraficamente desde 1891 (quando o Carbonífero foi dividido). O Período Ediacaran é definido como o período entre 630 milhões de anos atrás e o início do Cambriano, cerca de 542 milhões de anos atrás (Knoll, 2004).
Quão significativas são estas datas? Até há pouco tempo, pensava-se que uma diversidade de animais complexos apareceu subitamente na “explosão do Cambriano”, que começou há cerca de 545 milhões de anos atrás. Sabemos agora que os animais existiam pelo menos 60 milhões de anos antes disto e talvez até 600 milhões de anos antes do Cambriano. Sessenta milhões de anos podem não parecer tão longos, dadas as vastas extensões de tempo no Pré-Cambriano, mas uma grande parte da evolução animal pode ocorrer em tal período. Foram necessários cerca de sessenta milhões de anos para que o peixe rhipidistiano evoluísse para os primeiros anfíbios e cerca de 60 milhões de anos para a subsequente diversificação destes anfíbios numa variedade de linhagens, incluindo os primeiros répteis. Um período de sessenta milhões de anos com início no final do Permiano incluiria duas extinções em massa. Começaria com um mundo dominado por répteis sinápticos e os arcaicos primitivos terminariam no Jurássico com as suas diversas linhagens de dinossauros, pterossauros, e répteis marinhos. Em cerca de sessenta milhões de anos, os mamíferos primitivos que sobreviveram à extinção dos dinossauros irradiaram para os diversos grupos fósseis e vivos do Cenozóico.


2) SÃO ANIMAIS SIMPLES
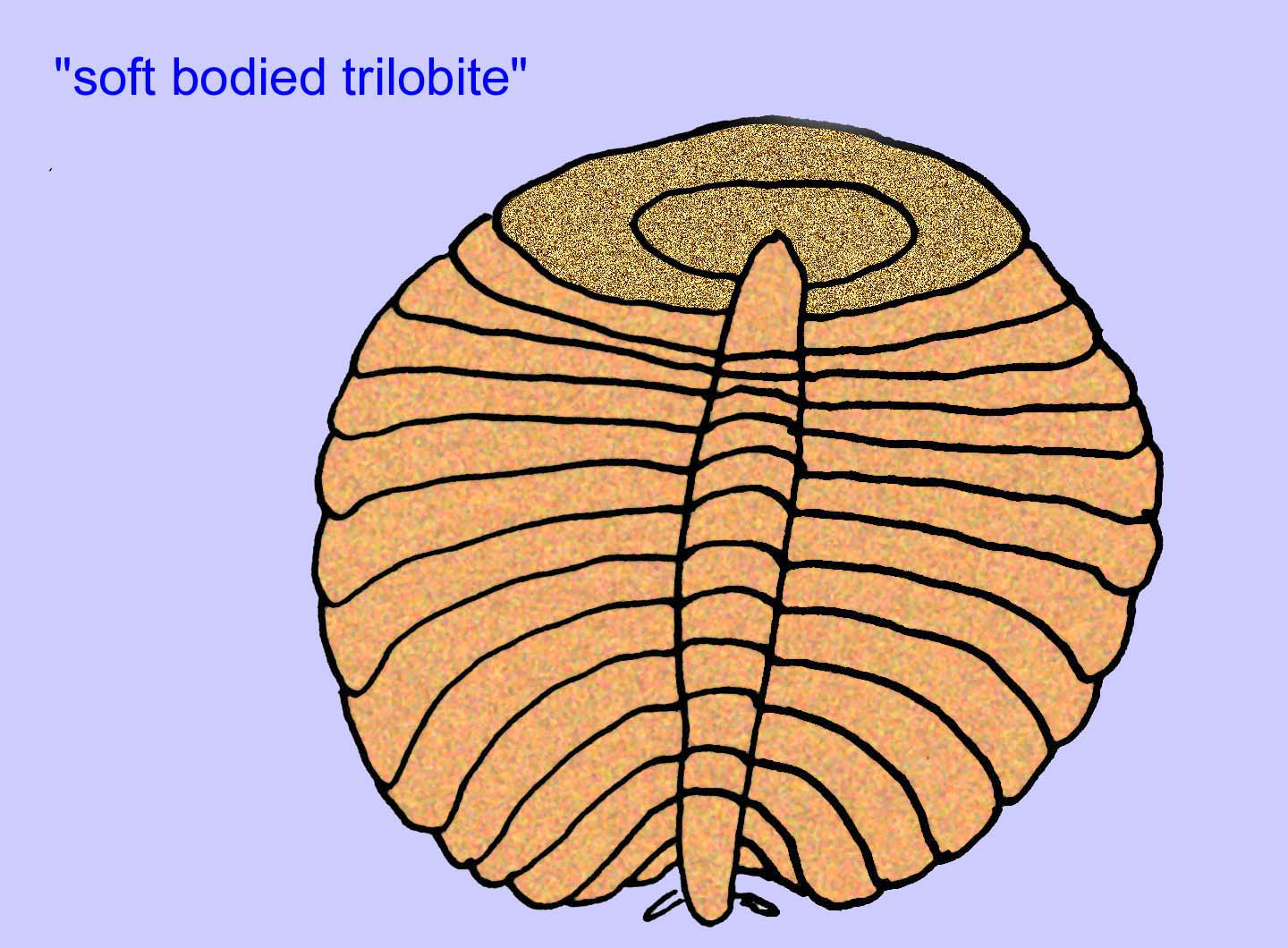
A maioria dos organismos Ediacaran têm menos de uma polegada de tamanho, mas alguns têm vários pés de comprimento. São estranhos e a maioria são muito diferentes de qualquer coisa viva hoje em dia (muitos referem-se a eles como o grupo Vendobionta). Praticamente todos não têm cabeça ou cauda, interiores/exteriores, frontais/trastras, dentes, ou olhos. A maioria não tem sistemas circulatórios, nervosos, ou digestivos óbvios, nem órgãos internos de qualquer tipo. A maioria é muito fina e plana – uma condição que seria essencial para animais sem sistemas circulatório, respiratório, digestivo ou excretor. Alguns locais têm uma diversidade de animais, com até 30 taxas diferentes (Buss, 1994).
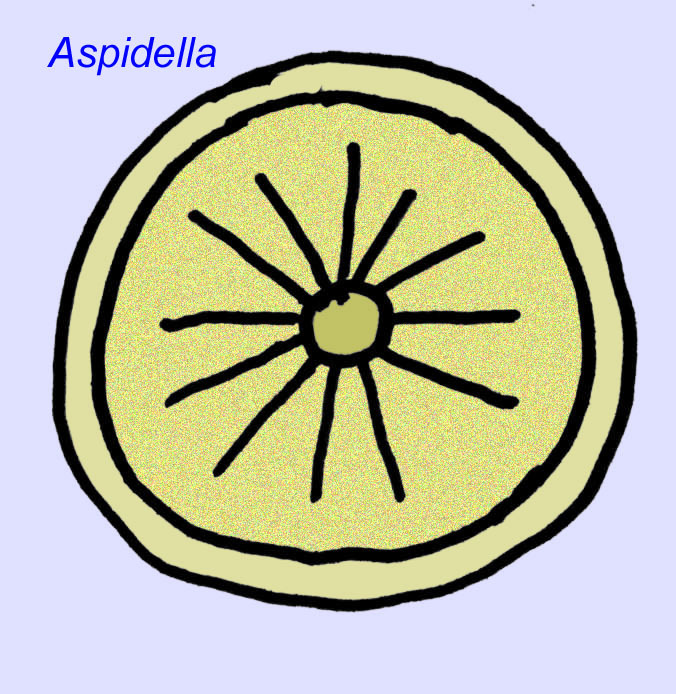
O que são eles? Houve uma diversidade de opiniões sobre estes organismos no passado: alguns classificaram-nos como plantas (a conclusão a partir da Primeira Guerra Mundial), organismos unicelulares muito grandes, líquenes únicos, um conjunto de animais completamente extinto, e um grupo de animais que inclui os antepassados dos animais modernos. Inicialmente pensava-se que tinham sido extintos muito antes da ascensão dos grupos de animais modernos, agora é óbvio que alguns grupos sobreviveram ao Cambriano. Alguns podem representar fósseis de animais não-metzóicos (Droser, 2006). A densidade de indivíduos do animal discoidal Aspidella indica que a biomassa de animais de fundo no Pré-Cambriano poderia ter-se aproximado da observada nas comunidades modernas (Gehling, 2000).
Muitos dos fósseis pré-Ediacaran representam simples colónias multicelulares de bactérias, algas eucarióticas (incluindo algas verdes e vermelhas), e formas que têm semelhanças com fungos. Alguns fósseis neoproterozóicos foram interpretados como espículas e larvas de esponjas (Butterfield, 2009).
Esponjas, que são um dos tipos de animais mais simples, foram identificados a partir de sedimentos Ediacaran (como o género Palaeophragmodictya), que são semelhantes a algumas esponjas Paleozoicas. Estas esponjas poderiam medir 1,5-10 cm de diâmetro (Gehling, 1996; Kerr, 1998a). As espículas de esponja estão presentes nas rochas de Ediacaran (Xiao, 2000).
alguns fósseis de Ediacaran têm sido interpretados como fungos e animais basais mais primitivos do que esponjas (Xiao, 2009).
CNIDARIANS
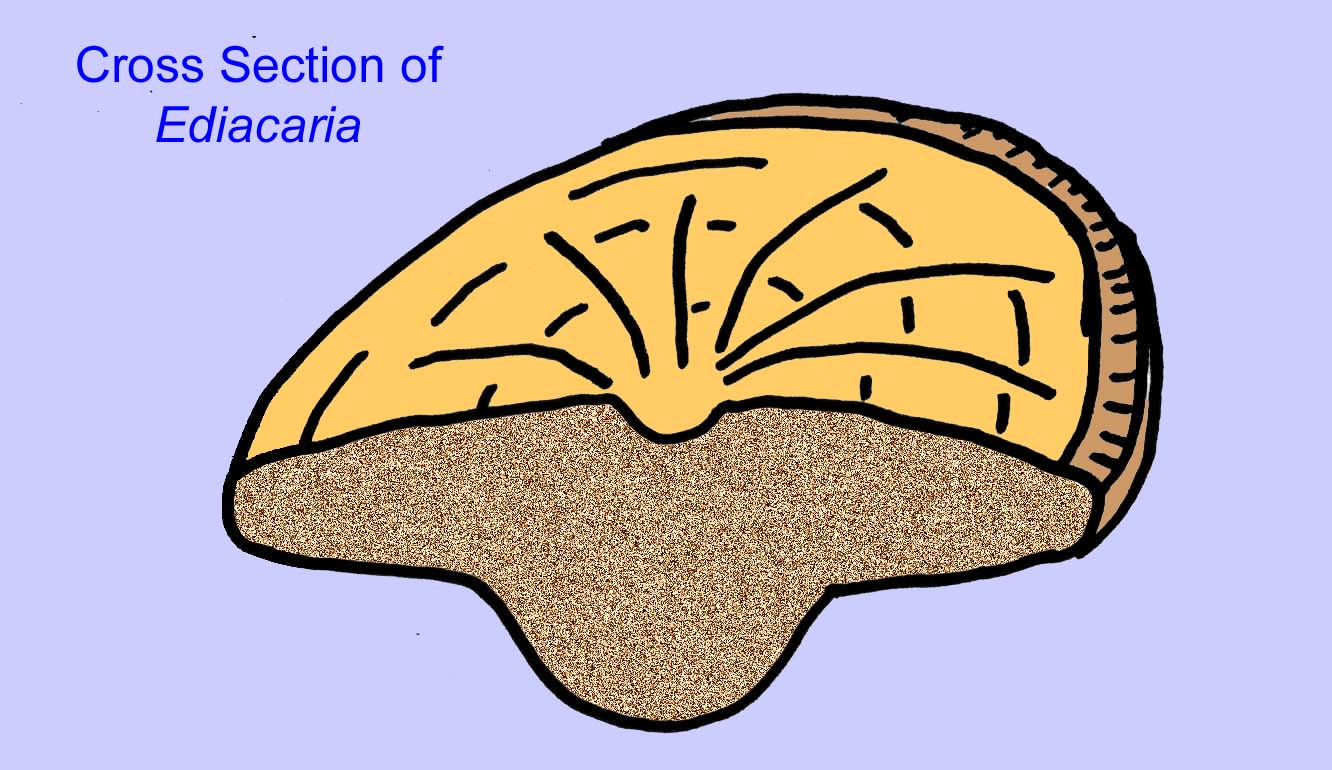
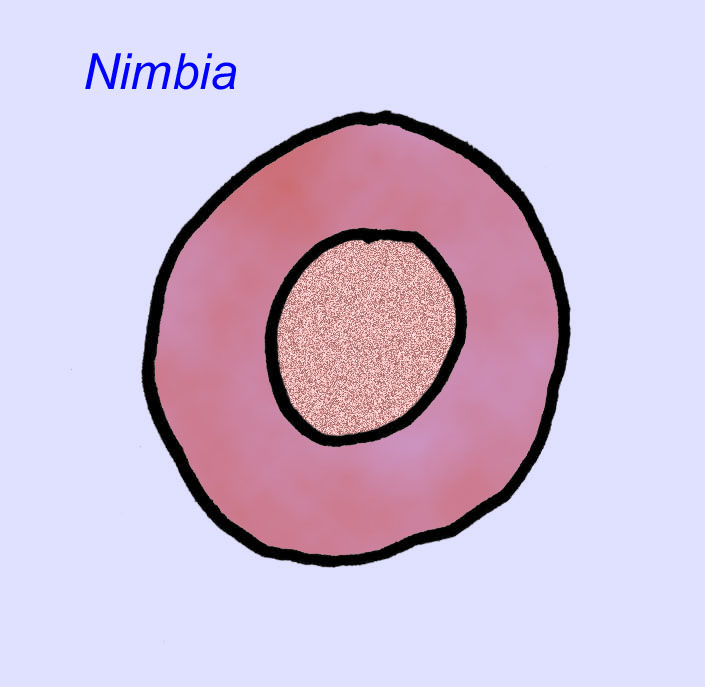
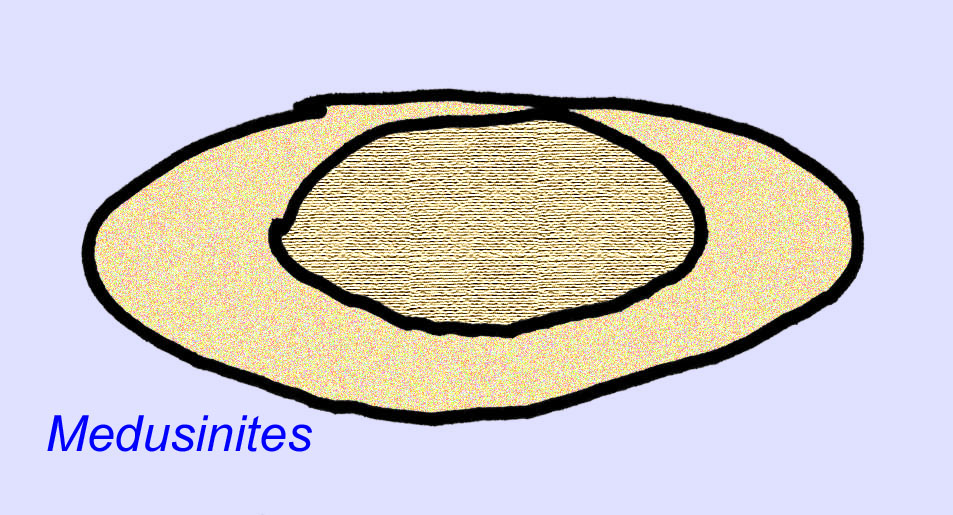
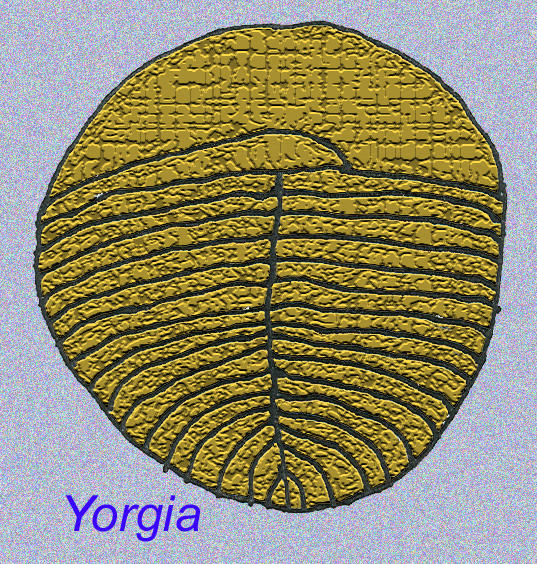
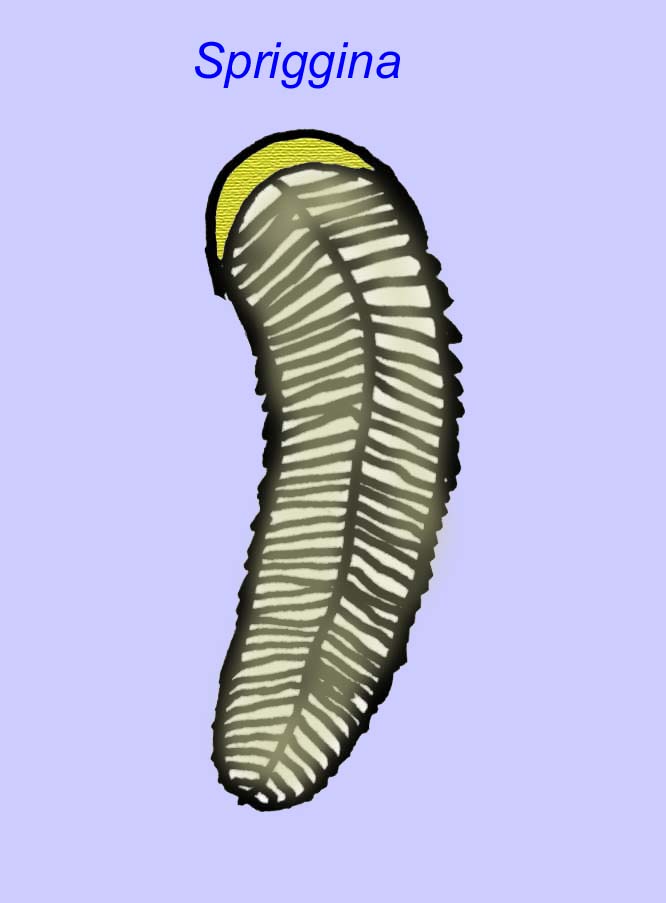
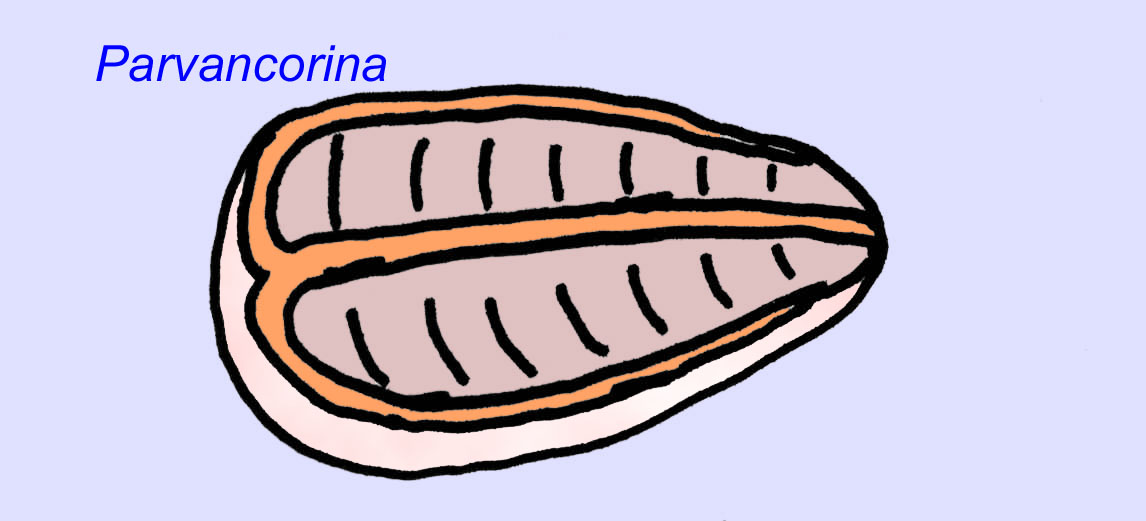
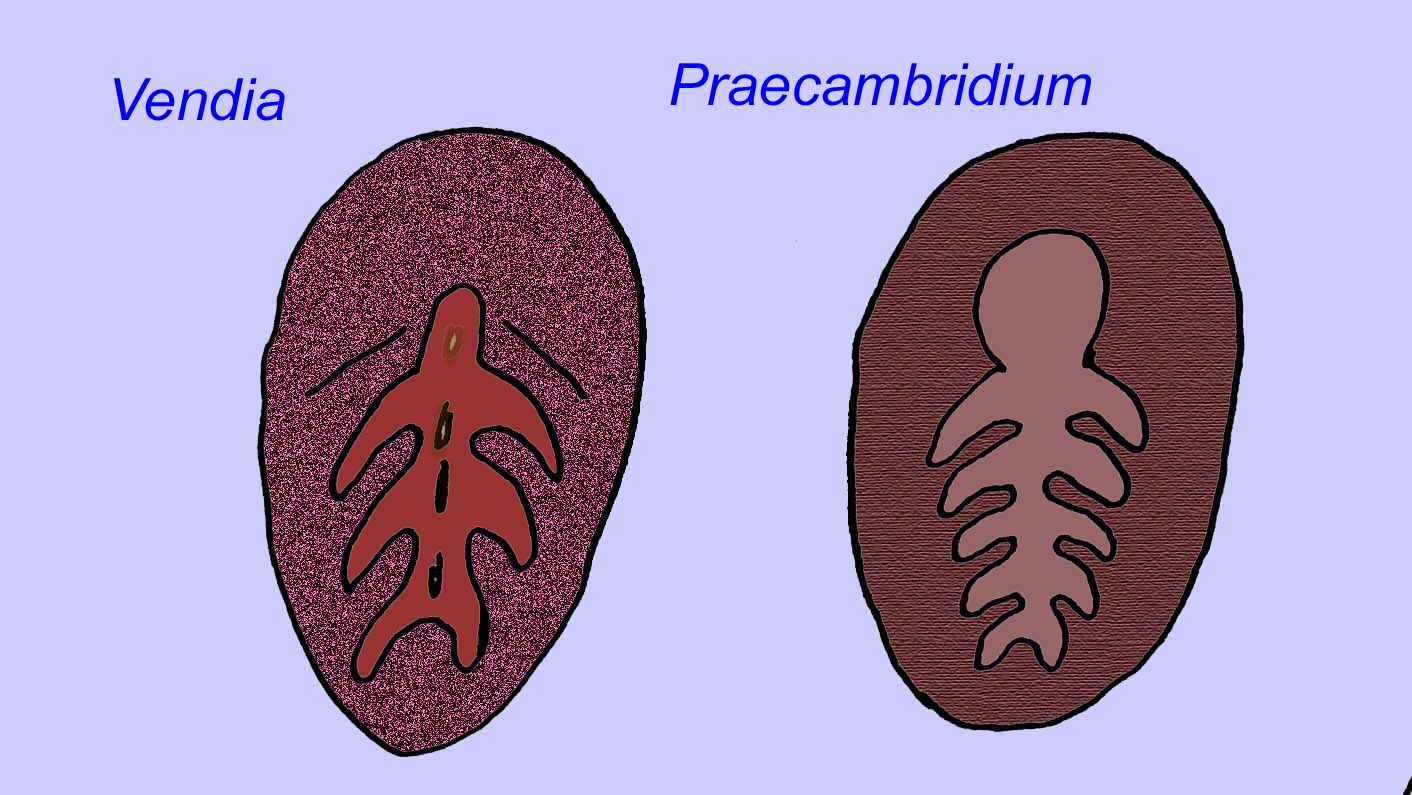
Há um consenso geral de que muitos fósseis Ediacaran devem ser classificados como primeiros cnidários, um grupo de animais que inclui as medusas modernas, os compartimentos marítimos, e os fãs do mar. Algumas formas (tais como Ediacaria e Nimbia) são radialmente simétricas e superficialmente semelhantes aos cnidários mas carecem de qualquer sinal de boca que possa ser uma razão para os excluir deste grupo. (Não é certo se os primeiros cnidários teriam tido bocas como todas as formas posteriores têm). Outros, tais como Albumares brunsae, possuem um sistema gastrovascular semelhante ao dos cnidários (Lipps, 1992; Sokolov, 1985). Muitos têm uma estrutura discoidal central num dos lados dos seus corpos planos e redondos (Crimes, 1995; Bertrand-Sarfati, 1995). Alguns animais Ediacaran (Spriggina, Parvancorina, e Praecambridium) têm distinções entre os seus lados superior e inferior (posterior e anterior). Alguns, como o Ediacaria, tinham linhas radiais e espessamento periférico (Crimes, 1995). Alguns dos cnidários de Ediacaran foram classificados como uma subclasse separada mas extinta (McCall, 2006).
enquanto a estes animais faltava um esqueleto mineralizado, a maioria ou mesmo todos (como Cyclomedusa e Ernietta) possuíam algum tipo de esqueleto rígido que era capaz de causar uma impressão na lama. O tipo de tecido que compõe o esqueleto interno não é conhecido (Crimes, 1995; Dzik, 1999).
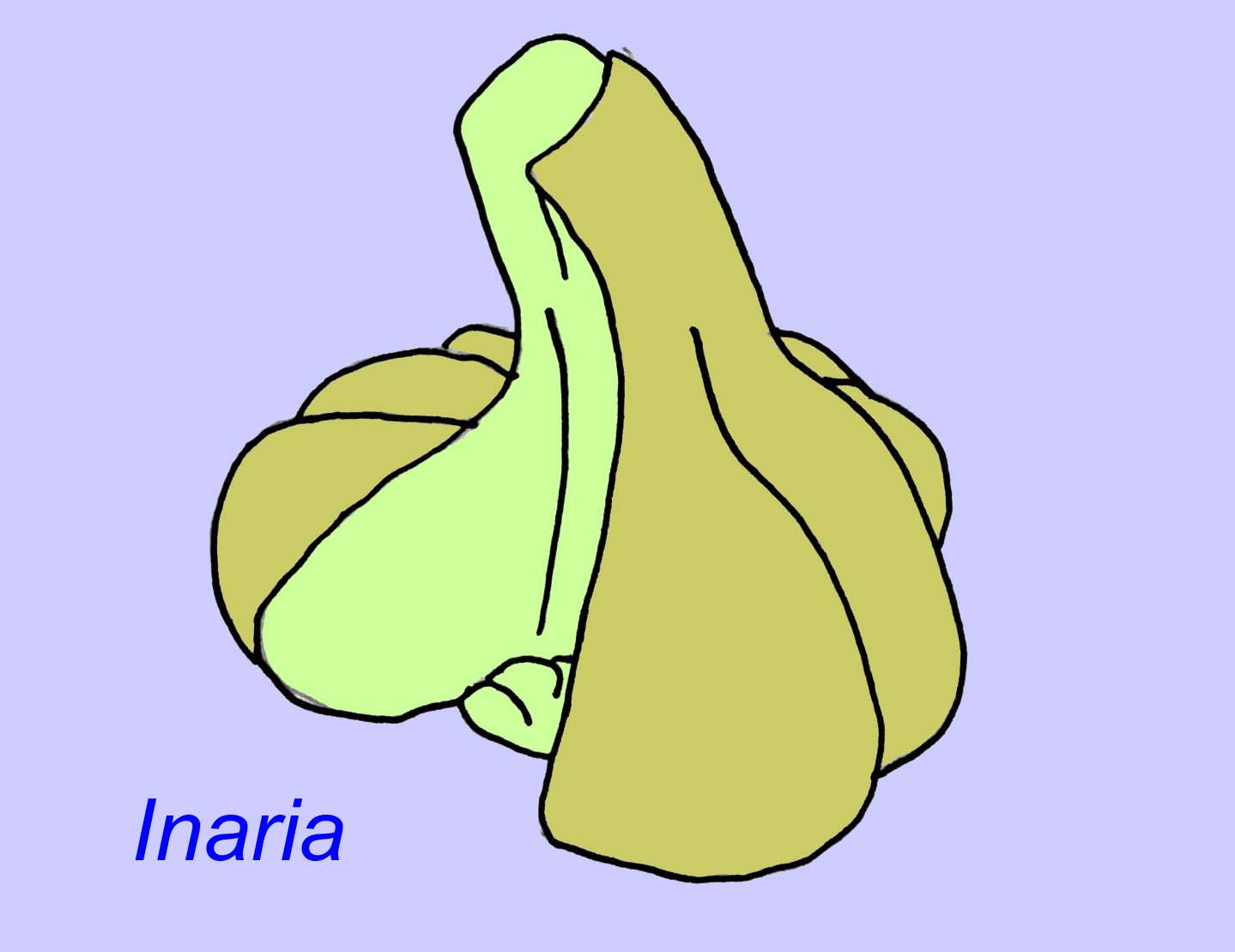
Mackenzia também parece ser um cnidário (mesmo que não seja, é quase certamente um animal metazoário). Há provas indirectas de bolus alimentares numa cavidade digestiva e de musculatura (Morris, 1993a). Sinocyclocyclicus pode ser um cnidário basal (ou mesmo um metazoário basal) cujo desenvolvimento apical é semelhante ao de alguns cnidários (como os corais tabulados) (Xiao, 2000). Inaria era um cnidário da classe Anthozoa com uma borda de boca, uma faringe, e uma câmara que pode ter hospedado organismos fotossintetizadores como os encontrados em muitos corais modernos (McMenaman, 1990, Gehling, 1988).
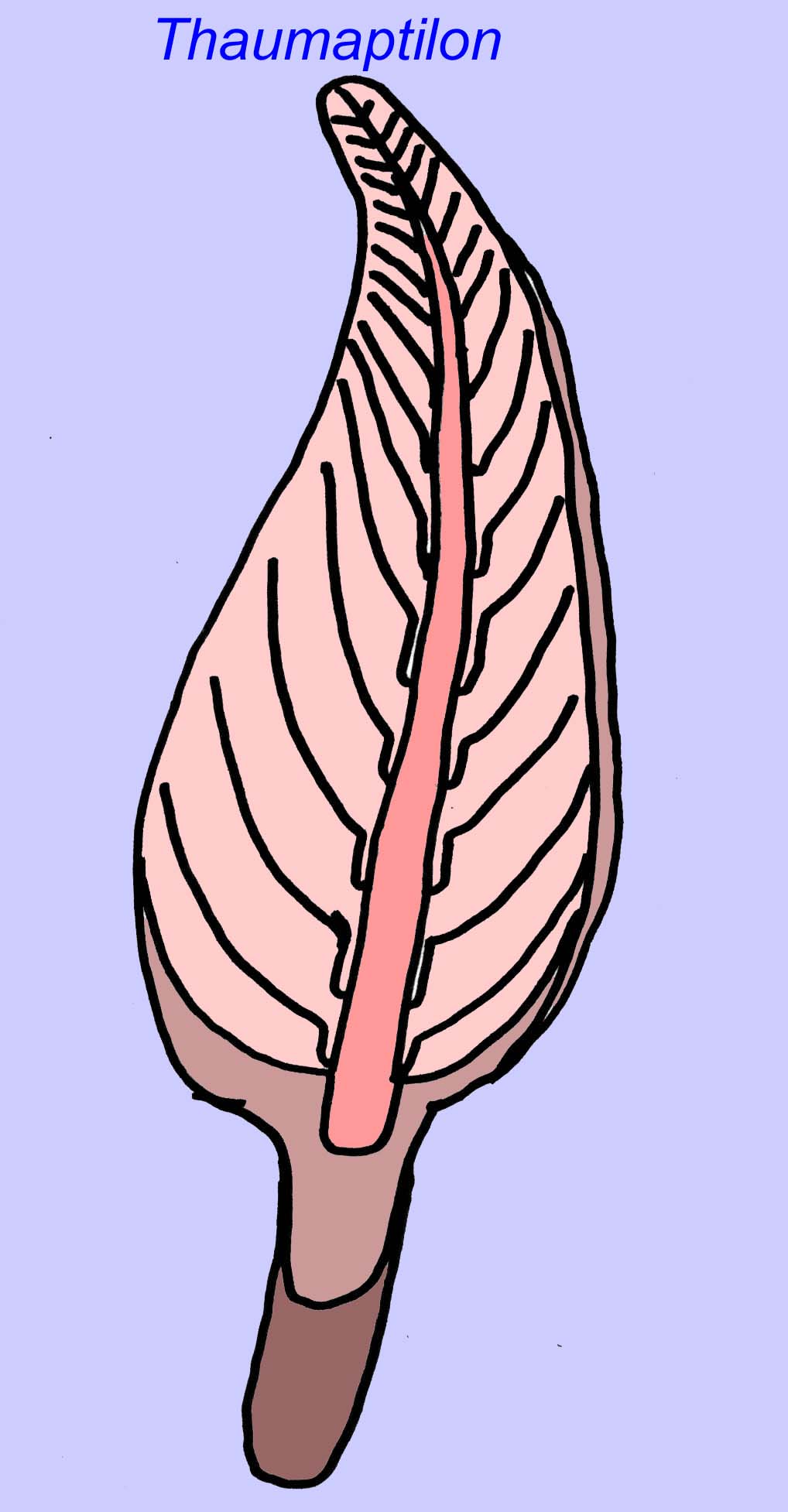
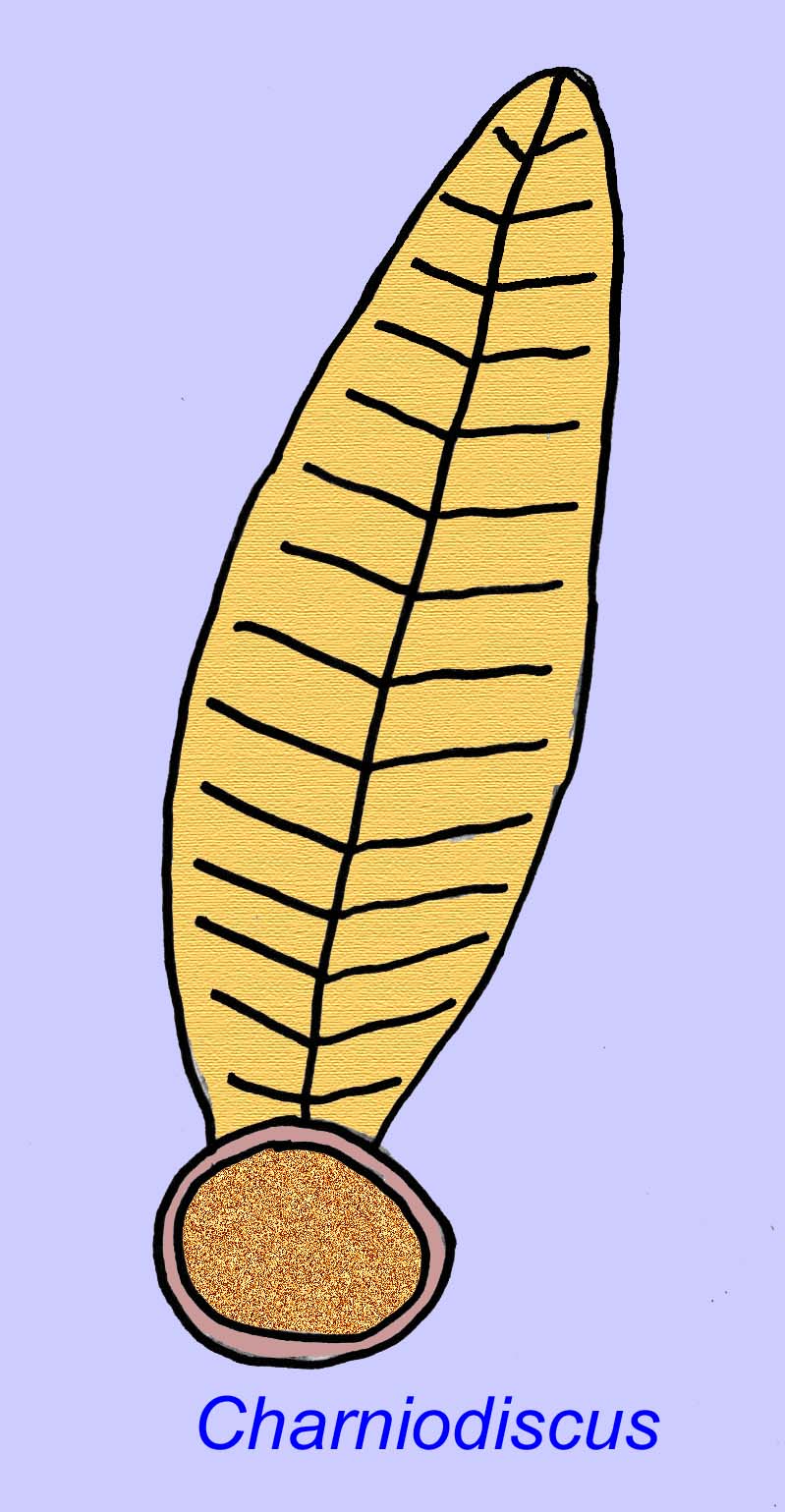
Thaumaptilon era um animal semelhante a um animal da frente que parece estar relacionado com os currais marinhos modernos. Ter-se-ia alimentado de detritos particulados e utilizado um holdfast para se fixar ao fundo (Morris, 1993a; 1993b). O frond de Charniodiscus podia atingir um comprimento de 1,2 metros (Sokolov, 1990).
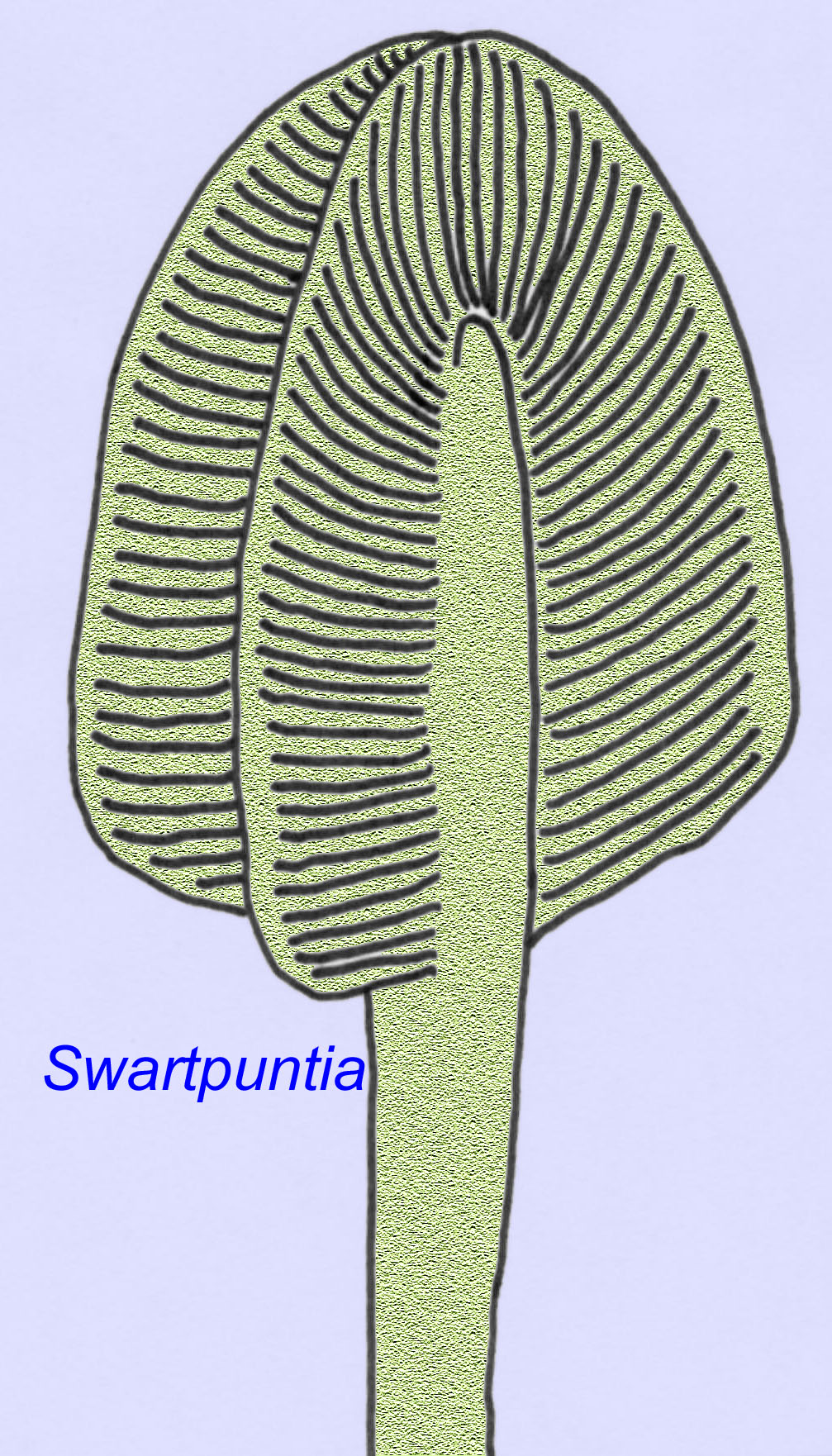
Muitos animais Ediacaran (tais como Pteridinium, Swartpuntia, Charnia, e Charniodiscus) possuem um talo erecto unido a petaloides segmentados.
Um exemplar de Charnodiscus media 1,2 metros de comprimento.
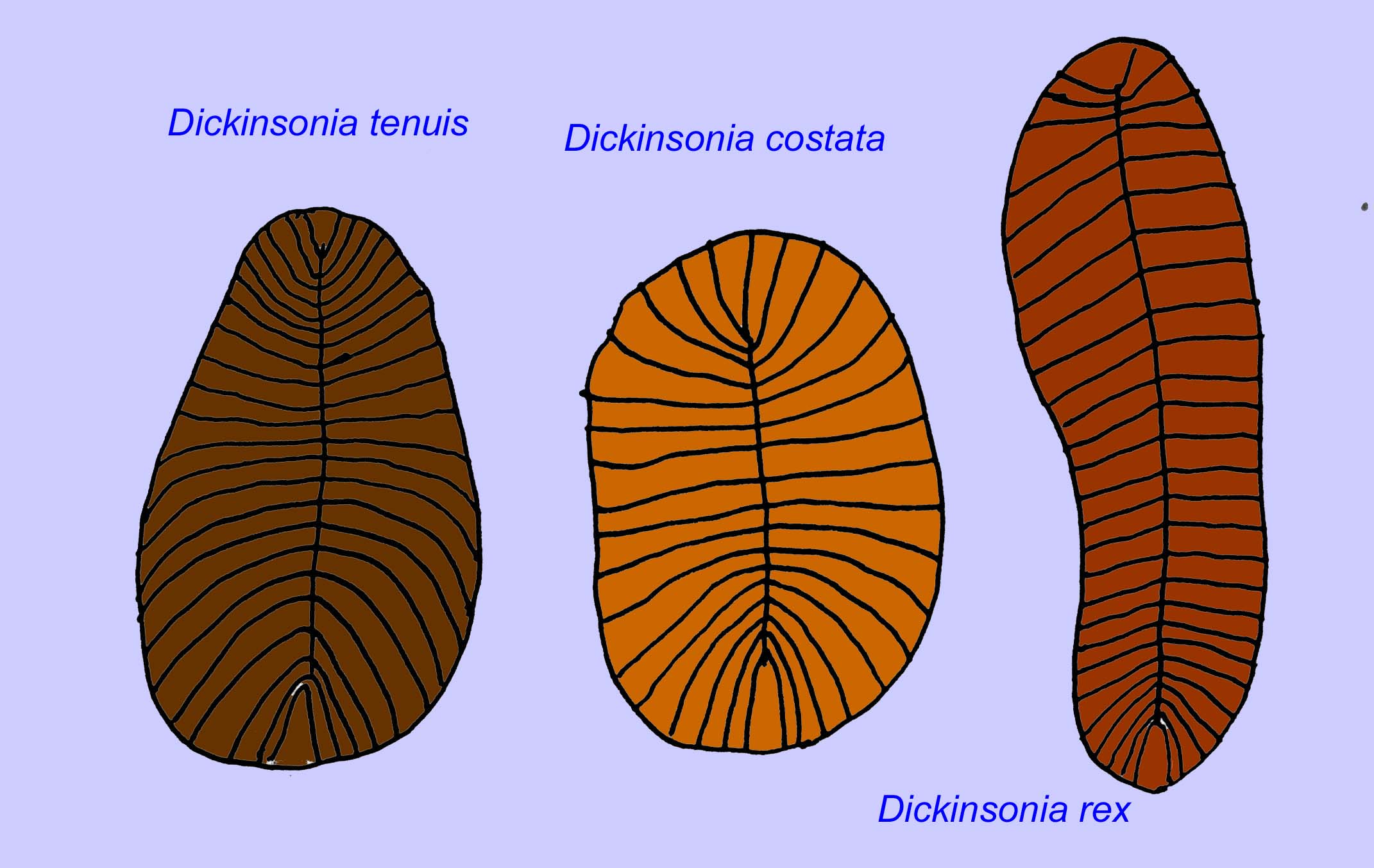
A anatomia microscópica de Swartpuntia é muito semelhante à de Dickinsonia, um verme precoce (Narbonne, 1997; McCall, 2006). Os embriões fossilizados que parecem pertencer a animais bilateranos são conhecidos de há 570 milhões de anos (Xiao, 1998).

WORMS
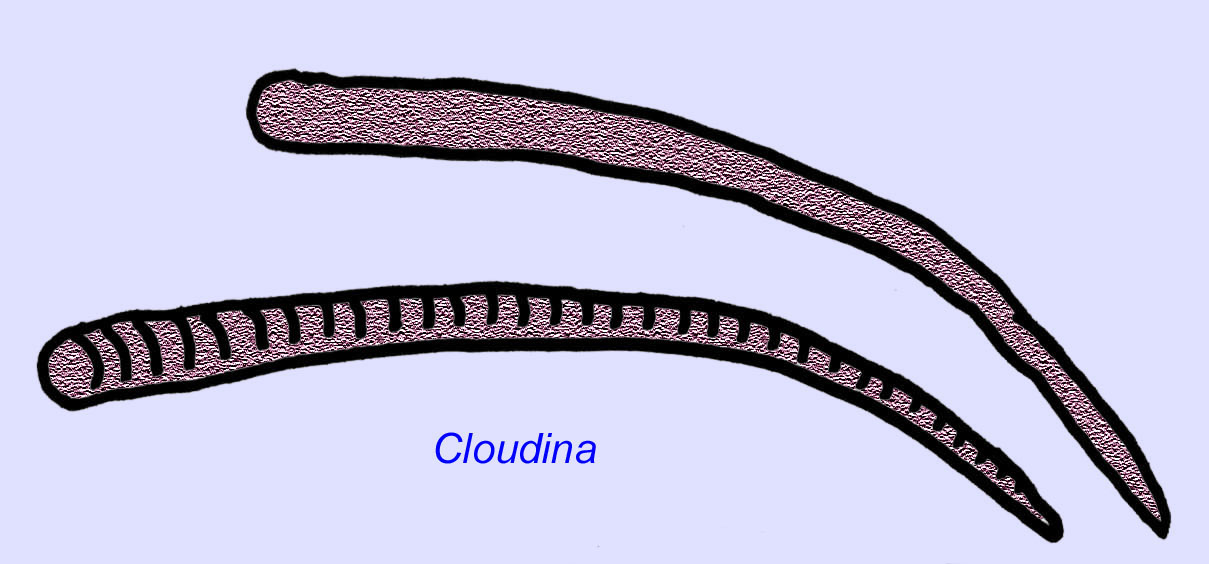
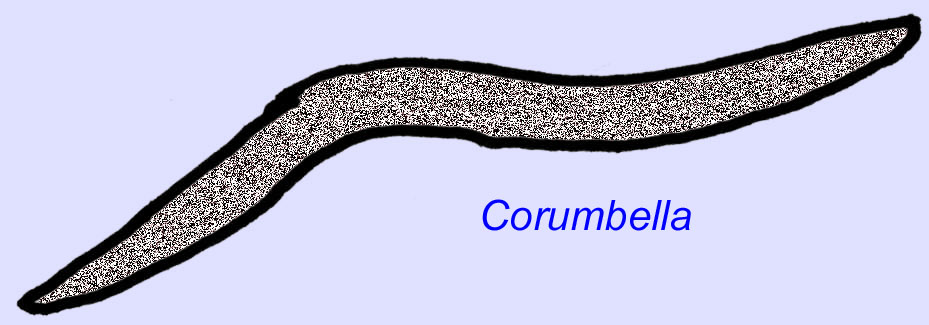
Vermes Ediacaran são conhecidos da Ásia, Europa, África, Austrália, e América do Norte (McCall, 2006). Vários animais Ediacaran eram tubulares como Archaeichnium, Cloudina, Corumbella, e Onuphionella. Enquanto Onuphionella foi comparada a vermes anelídeos, Corumbella foi comparada aos tubos construídos pelos pólipos de alguns cnidários. Enquanto Archaeichnium era flexível, Cloudina era coberta por um esqueleto mineralizado. Alguns tubos de Cloudina (comprimento máximo inferior a 4 cm) foram anulados e outros eram lisos, embora isto possa reflectir diferenças na preservação (Hagadorn, 2000). A Cloudina tem o primeiro animal conhecido com um esqueleto mineralizado e 3% dos espécimes têm buracos que indicam uma predação. Pode ser que esta predação tenha encorajado as estruturas mais duras típicas de muitos dos animais do Cambriano (Bengston, 1992). Cloudina é conhecida do Pré-Cambriano da América do Sul, Europa, Médio Oriente e Sudeste Asiático e os seus parentes são conhecidos do Cambriano (antes da disseminação dos trilobitas) na América do Norte (Grant, 1990). Os fósseis de Cloudina são conhecidos desde pelo menos 6 milhões de anos antes do início do Período Cambriano (Condon, 2005).
Além das duas espécies reconhecidas de Cloudina, várias outras espécies fósseis possuíam conchas calcificadas no Pré-Cambriano tardio (McCall, 2006). Cloudina, Namacalanthus, e Conotubus são animais do Pré-Cambriano esqueletizado que também são conhecidos no Cambriano Primitivo (Shu, 2008).
Foram encontrados vários vermes como Paleolina evenkiana e Planolites beverlyensis no final do Proterozóico e no período Cambriano. Os fósseis de planólitos têm 1-2 mm de largura e 5 a 13 mm de comprimento (Bartley, 1998). Fósseis não segmentados tais como Protechirus, Platypholina, e Vladimissa não são bem compreendidos. Alguns dos fósseis são segmentados, tais como Dickinsonia, que é classificado como um verme anelídeo. Os espécimes mais pequenos de Dickinsonia têm 4 mm de diâmetro, enquanto que os maiores têm 1 metro de comprimento. Apesar do comprimento destes vermes, eles eram muito finos com menos de 3 mm. As evidências sugerem que a quantidade de oxigénio na atmosfera no final do Pré-Cambriano era apenas cerca de um décimo da sua concentração actual e a extrema magreza (e portanto grande área superficial) destes vermes pode ter sido uma adaptação para a difusão de oxigénio. Estes vermes eram comuns e generalizados. Alimentavam-se de detritos no fundo do oceano, dado o sedimento que frequentemente enchia o seu intestino (Runnegar, 1982). A natureza simples do Dickinsonia pode indicar que se trata de um cnidário e não de um bilateran (Brasier, 2008).
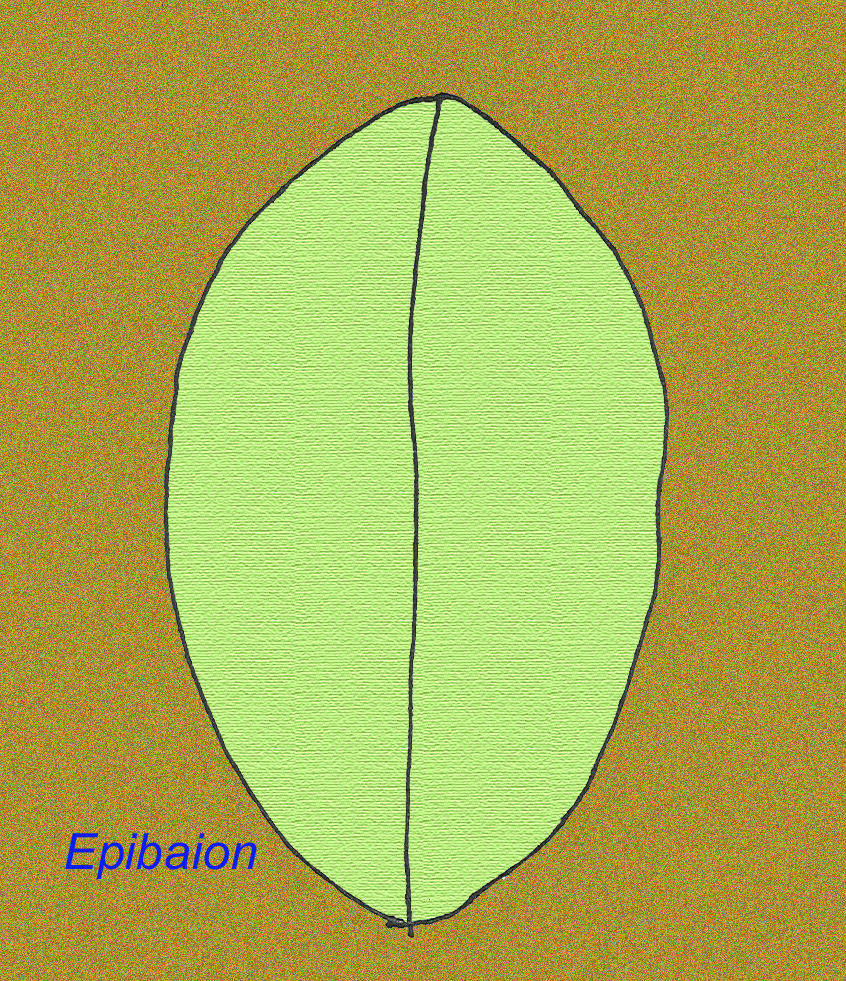
Um grande animal Ediacran chamado Epibaion axiferus deixou impressões reagindo até 4,3 metros por 0,15 metros (com um segundo espécime medindo 2,5 metros por 0,35 metros) (Ivantsov, 2002).


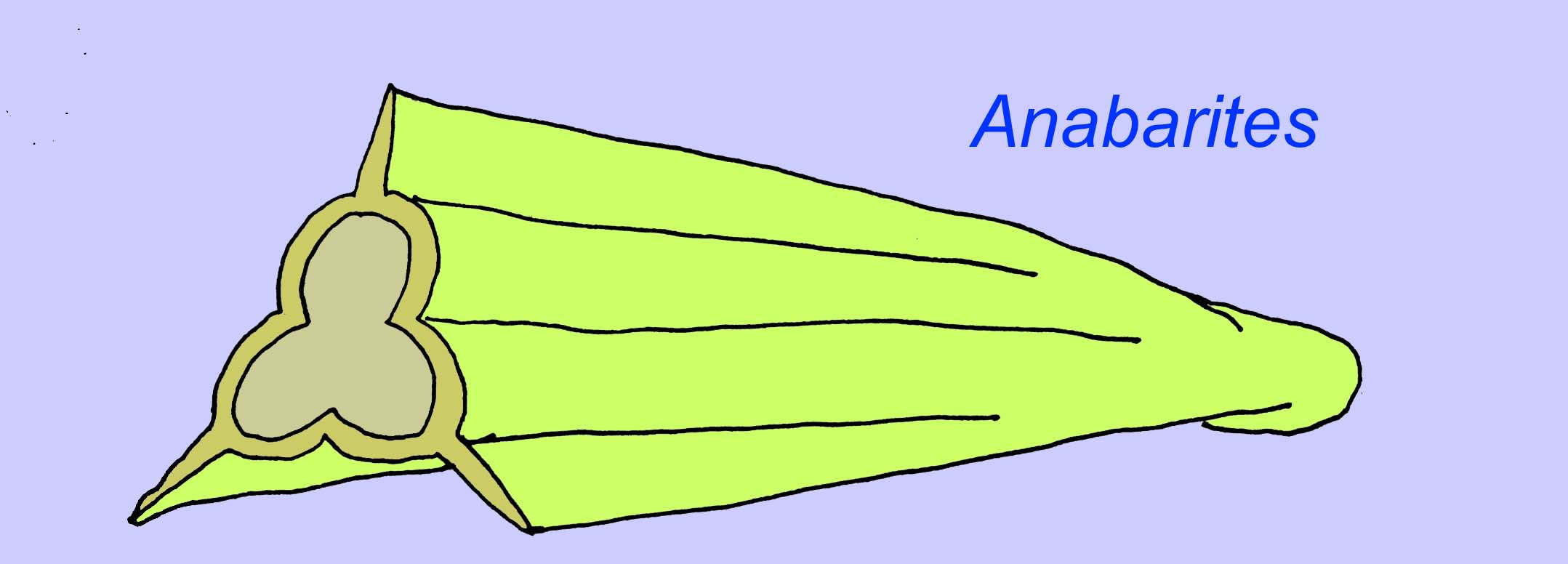
Anabaritids are shelled fossils of the early Cambrian. Alguns, como os anabaritas podem datar do fim do Pré-Cambriano (McMenamin, 1990).
Foram encontrados fósseis de tocas de minhocas em vários locais Proterozóicos. Estas tocas foram feitas por minhocas mais avançadas do que as minhocas planas. (Bartley, 1998, Narbonne, 1997, Hagadorn, 1998, McMenamin, 1996). Alguns cnidários modernos podem formar tocas simples e é possível que alguns dos vestígios de fósseis das tocas tenham sido formados por cnidários em vez de bilateranos (Jensen, 2005). Algumas tocas de vermes estão completas com excrementos, o que implica um sistema digestivo como o encontrado em vermes mais avançados (Morris, 1993a; Valentine 1994). Foram encontrados fósseis de embriões de vermes desde o Cambriano mais antigo (Bengston, 1997).
AREIS QUE OS ANIMAIS SÃO MAIS AVANÇADOS DO QUE OS PALAVRAS AMBORA A FAUNA EDIACARANA?
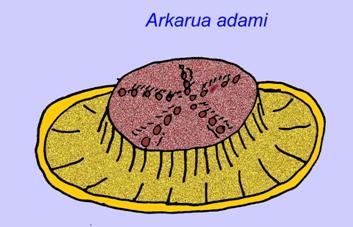
algumas análises genéticas sugeriram que a separação dos principais grupos de animais celómatos ocorreu antes do Cambriano (Gu, 1998). Vários fósseis de Ediacaran foram classificados em grupos de coelomatos, fornecendo apoio a esta conclusão. O Arkurua tem semelhanças com os equinodermos com a sua simetria de 5 ramos e as suas semelhanças com os equinodermos primitivos do Cambriano (os edrioasteroids). Talvez as 5 estruturas estejam a alimentar braços que levam a uma ranhura central (Gehling, 1987). Dois géneros (Redkinia e Spriggina) podem estar próximos da base de linhagens de artrópodes. Tribrachidium possuía três apêndices numa das suas superfícies foi encontrado na Austrália e na Europa (Wright, 1997b; McCall, 2006). Algumas da fauna Ediacaran tinham um plano corporal que era triradial (Tribrachidium, Ventagyrus), tetraradial (Conomedusies), e octaradial (Eoandromeda). Alguns interpretaram-nos como linhagens extintas de cnidários (Xiao, 2009).
O Ausia fóssil Ediacaran foi classificado como urochordate (Xiao, 2009).
Duas espécies, Phylozoon, e Kimberella (ver abaixo), podem ser parentes de vermes segmentados (annelídeos) ou moluscos. A Kimberella tem simetria bilateral e uma possível tripa (Fedonkin, 1997). A Kimberella parece ter sido um molusco que viveu como um predador marinho nadador rápido. Os buracos furados nas conchas de Cloudina foram atribuídos a predadores de moluscos Edicaran (McCall, 2006). Fósseis de embriões do Pré-Cambriano tardio formaram corpos polares semelhantes a muitos moluscos (Chen, 2006).
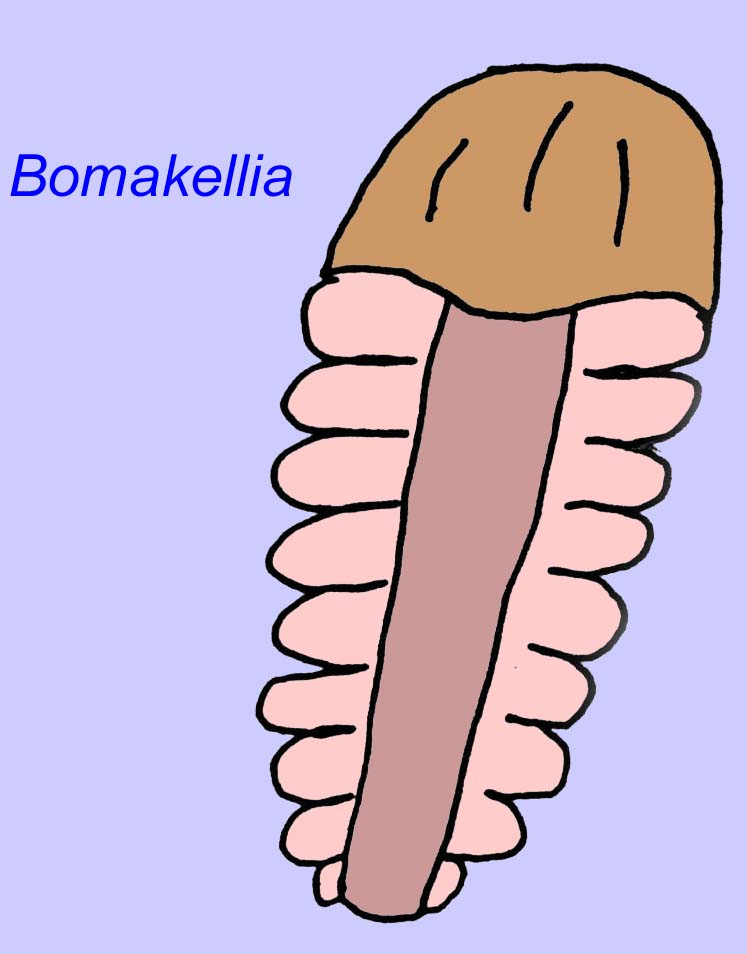
Outros fósseis podem estar relacionados com artrópodes, alguns dos quais (como Bomakellia) têm uma estrutura que tem sido comparada ao escudo da cabeça dos trilobitas (um tipo comum de artrópode da Era Paleozóica; McCall, 2006).
A fauna de Ediacaran tinha um plano corporal que era triradial (Tribrachidium, Ventagyrus), tetraradial (Conomedusies), e octaradial (Eoandromeda). Alguns interpretaram-nos como linhagens extintas de cnidários (Xiao, 2009).