Pierwsze zwierzęta
Najprymitywniejsze żywe istoty są jednokomórkowe: cały organizm składa się z jednej komórki. Niektóre organizmy mogą żyć jako pojedyncze komórki lub jako grupy komórek w kolonii. Jeśli komórki kolonii zostaną rozdzielone, mogą kontynuować życie jako pojedyncze komórki lub utworzyć nową kolonię. Żadne komórki nie są wyspecjalizowane do wykonywania konkretnych zadań. Współczesna alga Volvox jest tego dobrym przykładem: sąsiednie zdjęcie przedstawia kolonię Volvox.
Wielokomórkowość ewoluowała niezależnie kilkadziesiąt razy w roślinach, algach czerwonych, algach brunatnych, grzybach i zwierzętach. Na przykład, glony czerwone w grupie Volvocales obejmują gatunki jednokomórkowe (Chlamydomonas), uogólnione gatunki czterokomórkowe (Basichlamys) i gatunki tworzące kolonie, które mogą obejmować 50 000 komórek kolonii (Volvox) (Sachs, 2008).
Podczas gdy wiele alg żyje w tych prostych koloniach, niektóre algi rzeczywiście mają komórki wyspecjalizowane do wykonywania różnych zadań (mają tkanki, które są podobne do korzeni w funkcji i podobne do liści w funkcji).
Pierwsze skamieniałości glonów znane są sprzed około 2,1 miliarda lat, a od około 1 miliarda lat obserwuje się wzrost różnorodności znanych gatunków glonów. Wydaje się, że glony eukariotyczne zróżnicowały się nagle około 600 milionów lat temu, po epokach lodowcowych końca proterozoiku (Cavalier-Smith, 2000).
Do 570 milionów lat temu znane są glony o różnych cechach anatomicznych i reprodukcyjnych (Xiao, 1998). W zapisie kopalnym zachowała się różnorodność prehistorycznych glonów, takich jak te przedstawione poniżej.
FUNGI
Wiele skamieniałości preediakarskich reprezentuje proste wielokomórkowe kolonie bakterii, eukariotyczne algi (w tym zielone i czerwone algi) oraz formy, które wykazują podobieństwo do grzybów. (Butterfield, 2009; Xiao, 2009).
W 100-milionowym bursztynie zaobserwowano mięsożerne grzyby tworzące pierścienie hyphalowe, żerujące na nicieniach (Schmidt, 2007).
PIERWSZE ZWIERZĘTA
Fauna ediakarska to zespół organizmów nazwanych tak od wzgórz Ediacara w Australii, gdzie zostały po raz pierwszy znalezione. Skamieniałości ediakarskie znane są z ponad 30 krajów i wszystkich kontynentów z wyjątkiem Antarktydy i Ameryki Południowej (De, 2005; McCall, 2006).Z fauny ediakarskiej opisano kilka tysięcy okazów, obejmujących ponad 270 gatunków, a wiele z nich nie zostało jeszcze opisanych (Fedonkin, 1990; Minnerop, 2000; Waggoner, 1998; Shen, 2008). Pierwsze ediakarskie zwierzę zostało opisane w 1872 roku, a drugie znalezisko w latach 1908-14 (McCall, 2006).
- THEY ARE OLD
Straty kopalne, które zostały datowane na więcej niż 1,2 miliarda lat zawierają zwierzęta – nie tylko zawierają skamieniałości promienisto-miękkich organizmów, ale zawierają nory, które wydają się być wykonane przez robaki produkujące śluz (Rasmussen, 2002; McCall, 2006). W Indiach znajduje się stanowisko, którego nory zostały datowane na 1,1 miliarda lat temu, choć niektórzy kwestionują tę datę. Ostatnio inne stanowisko z widocznymi śladami prymitywnych zwierząt (krewnych meduz) i norami robaków datowane jest na 1,2 mld lat (Seilacher, 1998; Kerr, 1998b; Morris, 1993). Inne skamieniałości ediakarskie znane są sprzed 610-600 mln lat, a bardziej zróżnicowane zespoły organizmów znane są sprzed 555,3 mln lat (McMenamin, 1996, Minnerop, 2000).Prekambryjskie warstwy skalne sprzed 580 mln lat posiadają skamieniałości małych bilateranów; większe zwierzęta bilateralne obecne są po datach sprzed 555 mln lat (Condon, 2005).Zostały one w dużej mierze wyparte przez zwierzęta „eksplozji kambryjskiej”, która rozpoczęła się około 545 mln lat temu. Okres ediakarski jest definiowany jako 635 milionów lat temu po zlodowaceniu Marinoan Snowball Earth (chociaż zlodowacenie Gaskiers 562 milionów lat temu wystąpiło w okresie ediakarskim) (Xiao, 2009).
Porównania molekularne współczesnych organizmów również potwierdzają wniosek, że zwierzęta wytworzyły wiele różnych linii przed kambrem. Na przykład jedno z badań 129 białek sugeruje, że eukarionty powstały między 950 a 1260 milionów lat temu, zwierzęta powstały między 761 a 957 milionów lat temu, a linie koelomatów rozdzieliły się między 642 a 761 milionów lat temu (Douzery, 2004). Inne analizy przyniosły podobne wyniki (Peterson, 2004). Inne molekularne szacunki pochodzenia zwierząt eumetazoan (cnidarians i bilaterans) dostarczone przez badania zegara molekularnego wahały się od 570 mln lat temu do 1100 mln lat temu. Pochodzenie zwierząt bilateralnych oszacowano na 580 milionów lat temu (Peterson, 2005). Skamieniałości zwierząt znalezione w Australii, Indiach i Teksasie datowane są na ponad 1 miliard lat, a segmentowane zwierzę z Rosji datowane jest na 890 milionów do 1 miliarda lat (De, 2005). Ze względu na ekscytujące odkrycia prekambryjskich zwierząt, do geologicznej skali czasu dodano nowy okres, który jest pierwszym okresem zdefiniowanym stratygraficznie od 1891 roku (kiedy to podzielono karbon). Okres ediakarski jest zdefiniowany jako czas pomiędzy 630 milionami lat temu a początkiem kambru, około 542 milionów lat temu (Knoll, 2004).
Jakie znaczenie mają te daty? Do niedawna sądzono, że różnorodność złożonych zwierząt pojawiła się nagle w „eksplozji kambryjskiej”, która rozpoczęła się około 545 milionów lat temu. Obecnie wiemy, że zwierzęta istniały przez co najmniej 60 milionów lat wcześniej, a być może nawet 600 milionów lat przed kambrem. Sześćdziesiąt milionów lat może nie wydawać się tak długim okresem, biorąc pod uwagę ogromne przestrzenie czasu w prekambrze, ale w takim czasie może nastąpić ogromna ewolucja zwierząt. Około sześćdziesięciu milionów lat zajęło rybom rhipidistycznym przekształcenie się w pierwsze płazy, a około 60 milionów lat późniejsze zróżnicowanie tych płazów w różne linie, w tym pierwsze gady. Okres sześćdziesięciu milionów lat rozpoczynający się pod koniec permu obejmowałby dwa masowe wymierania. Zaczęło się od świata zdominowanego przez gady synapsoidalne i prymitywne archozaury, a skończyło na Jurze z jej zróżnicowanymi liniami dinozaurów, pterozaurów i gadów morskich. W ciągu sześćdziesięciu milionów lat prymitywne ssaki, które przetrwały wyginięcie dinozaurów, przekształciły się w różnorodne kopalne i żywe grupy kenozoiku.


2) SĄ TO PROSTYCH ZWIERZĄT
Większość organizmów ediakarańskich ma rozmiar poniżej cala, ale niektóre mają kilka stóp długości. Są one dziwne i większość z nich bardzo różni się od wszystkiego, co żyje dzisiaj (wielu określa je jako grupę Vendobionta). Praktycznie wszystkie nie mają głowy ani ogona, wnętrza/zewnętrza, przodu/tyłu, zębów ani oczu. Większość z nich nie ma oczywistych układów krążenia, nerwowego, pokarmowego lub narządów wewnętrznych jakiegokolwiek rodzaju. Większość jest bardzo cienka i płaska – warunek, który byłby niezbędny dla zwierząt bez układu krążenia, oddechowego, trawiennego lub wydalniczego. Niektóre stanowiska charakteryzują się różnorodnością zwierząt, z aż 30 różnymi taksonami (Buss, 1994).
Czym one są? W przeszłości istniało wiele różnych opinii na temat tych organizmów: niektórzy klasyfikowali je jako rośliny (wniosek z I wojny światowej), bardzo duże organizmy jednokomórkowe, unikalne porosty, całkowicie wymarły zestaw zwierząt oraz grupę zwierząt, która obejmuje przodków współczesnych zwierząt. Pierwotnie sądzono, że wymarły one na długo przed powstaniem współczesnych grup zwierząt, obecnie jest oczywiste, że niektóre grupy przetrwały do kambru. Niektóre z nich mogą reprezentować skamieniałości zwierząt niemetazoanowych (Droser, 2006). Zagęszczenie osobników dyskoidalnego zwierzęcia Aspidella wskazuje, że biomasa zwierząt żyjących na dnie w prekambrze mogła zbliżyć się do tej obserwowanej we współczesnych społecznościach (Gehling, 2000).
Wiele przedindiakarskich skamieniałości reprezentuje proste wielokomórkowe kolonie bakterii, eukariotyczne glony (w tym zielone i czerwone) oraz formy przypominające grzyby. Niektóre skamieniałości neoproterozoiczne zostały zinterpretowane jako szkielety i larwy gąbek (Butterfield, 2009).
Gąbki, które są jednym z rodzajów najprostszych zwierząt, zostały zidentyfikowane z osadów ediakarskich (np. rodzaj Palaeophragmodictya), które są podobne do niektórych gąbek paleozoicznych. Gąbki te mogą osiągać średnicę 1,5-10 cm (Gehling, 1996; Kerr, 1998a). Spicule gąbek są obecne w skałach ediakarskich (Xiao, 2000).
Niektóre ediakarskie skamieniałości zostały zinterpretowane jako grzyby i zwierzęta bazalne, bardziej prymitywne od gąbek (Xiao, 2009).
CNIDARIANY
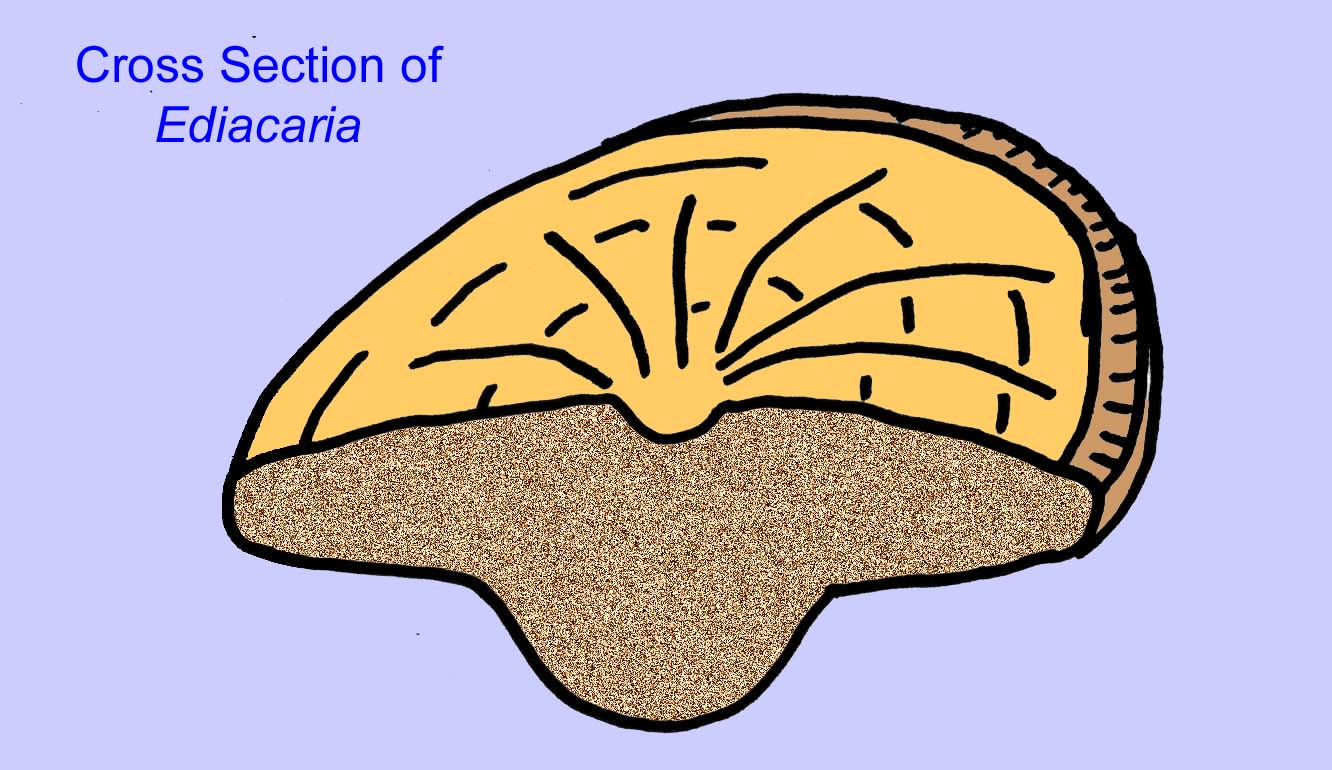
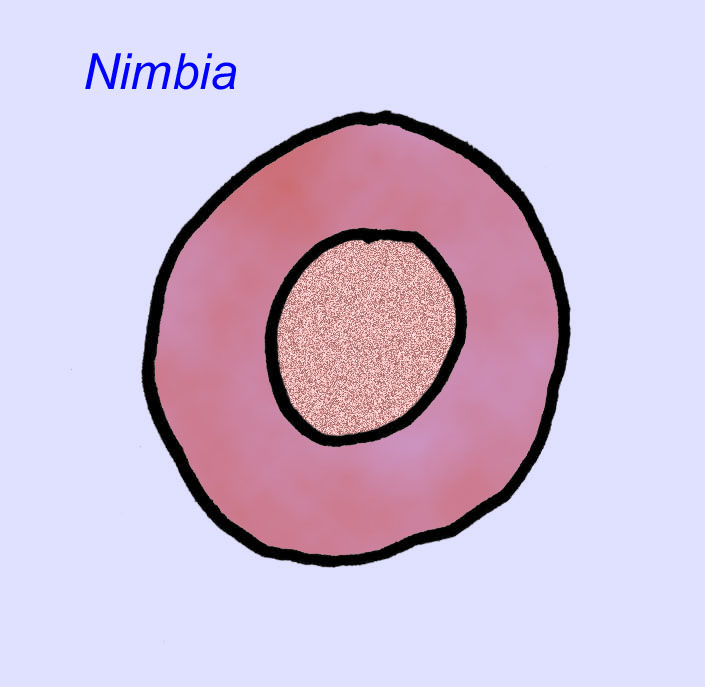
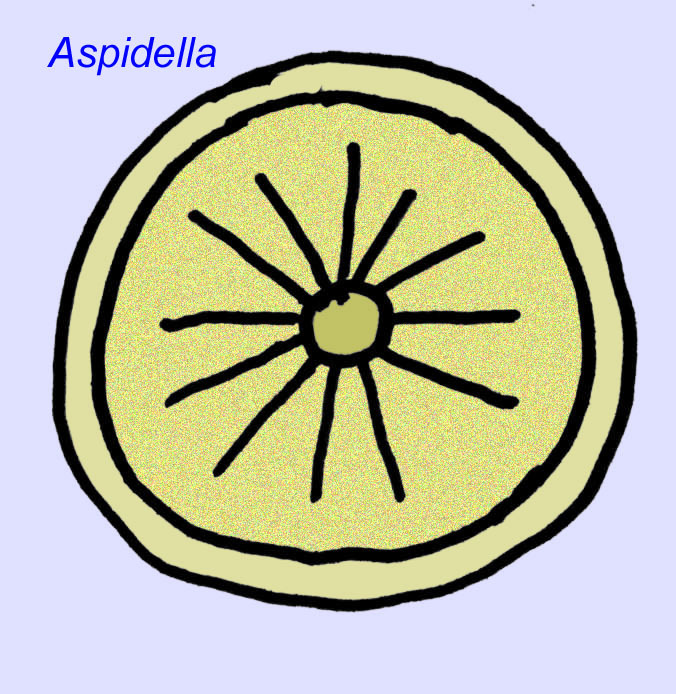
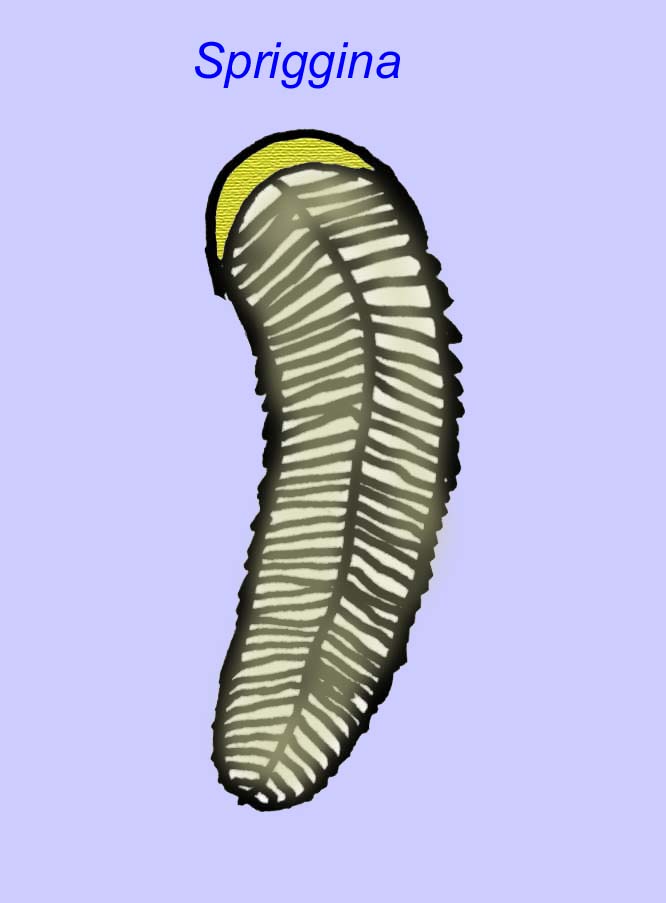
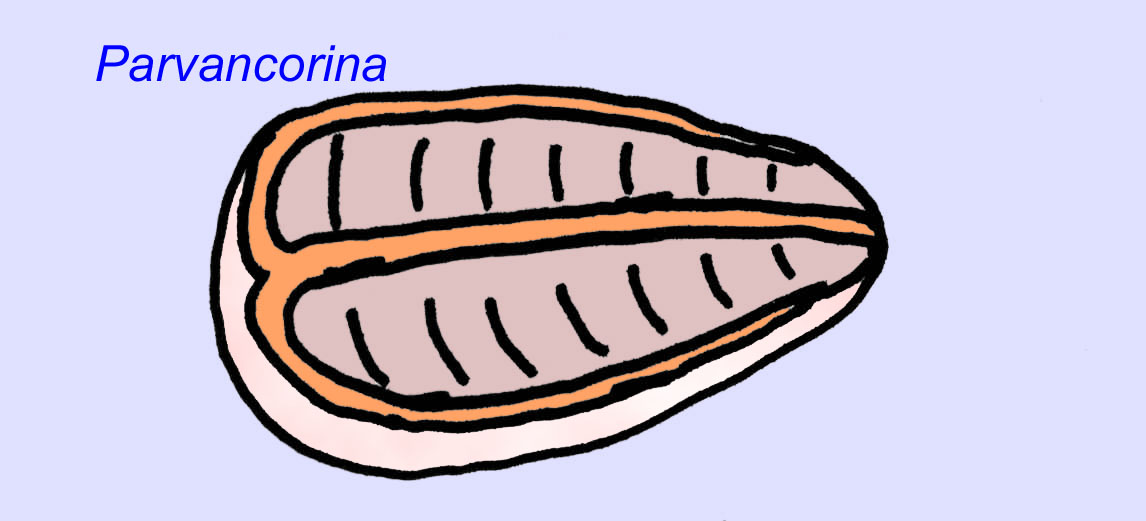
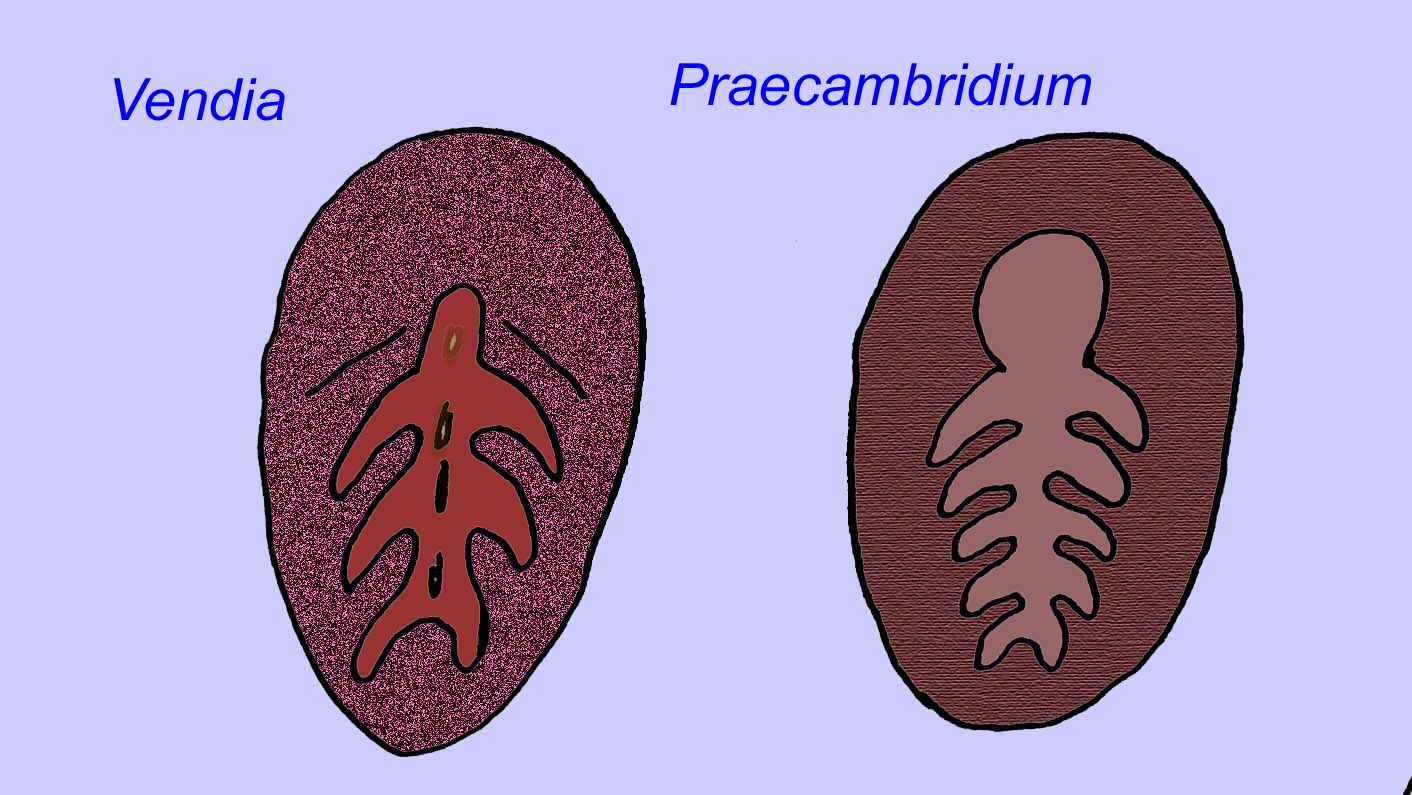
Istnieje powszechna zgoda co do tego, że wiele ediakarskich skamieniałości należy zaliczyć do wczesnych knidarian, grupy zwierząt, do której należą współczesne meduzy, pióra morskie i wachlarze morskie. Niektóre formy (takie jak Ediacaria i Nimbia) są promieniście symetryczne i powierzchownie podobne do knidriów, ale brak im jakichkolwiek oznak ust, co może być powodem do wykluczenia ich z tej grupy. (Nie jest pewne, czy pierwsze knidariony miałyby usta, tak jak wszystkie późniejsze formy). Inne, takie jak Albumares brunsae, posiadają układ pokarmowy podobny do tego, jaki posiadają knidaria (Lipps, 1992; Sokolov, 1985). Wiele z nich ma centralną strukturę dyskoidalną po jednej stronie płaskiego, okrągłego ciała (Crimes, 1995; Bertrand-Sarfati, 1995). Niektóre ediakarańskie zwierzęta (Spriggina, Parvancorina i Praecambridium) mają rozróżnienie między górną i dolną (tylną i przednią) stroną ciała. Niektóre, takie jak Ediacaria, miały linie promieniste i obwodowe zgrubienia (Crimes, 1995). Niektóre z ediakariańskich knidriów zostały sklasyfikowane jako odrębna, ale wymarła podklasa (McCall, 2006).
Pomimo że zwierzętom tym brakowało zmineralizowanego szkieletu, większość lub nawet wszystkie (jak Cyclomedusa i Ernietta) posiadały jakiś rodzaj sztywnego szkieletu, który był w stanie odcisnąć się w błocie. Rodzaj tkanki, z której zbudowany był wewnętrzny szkielet nie jest znany (Crimes, 1995; Dzik, 1999).
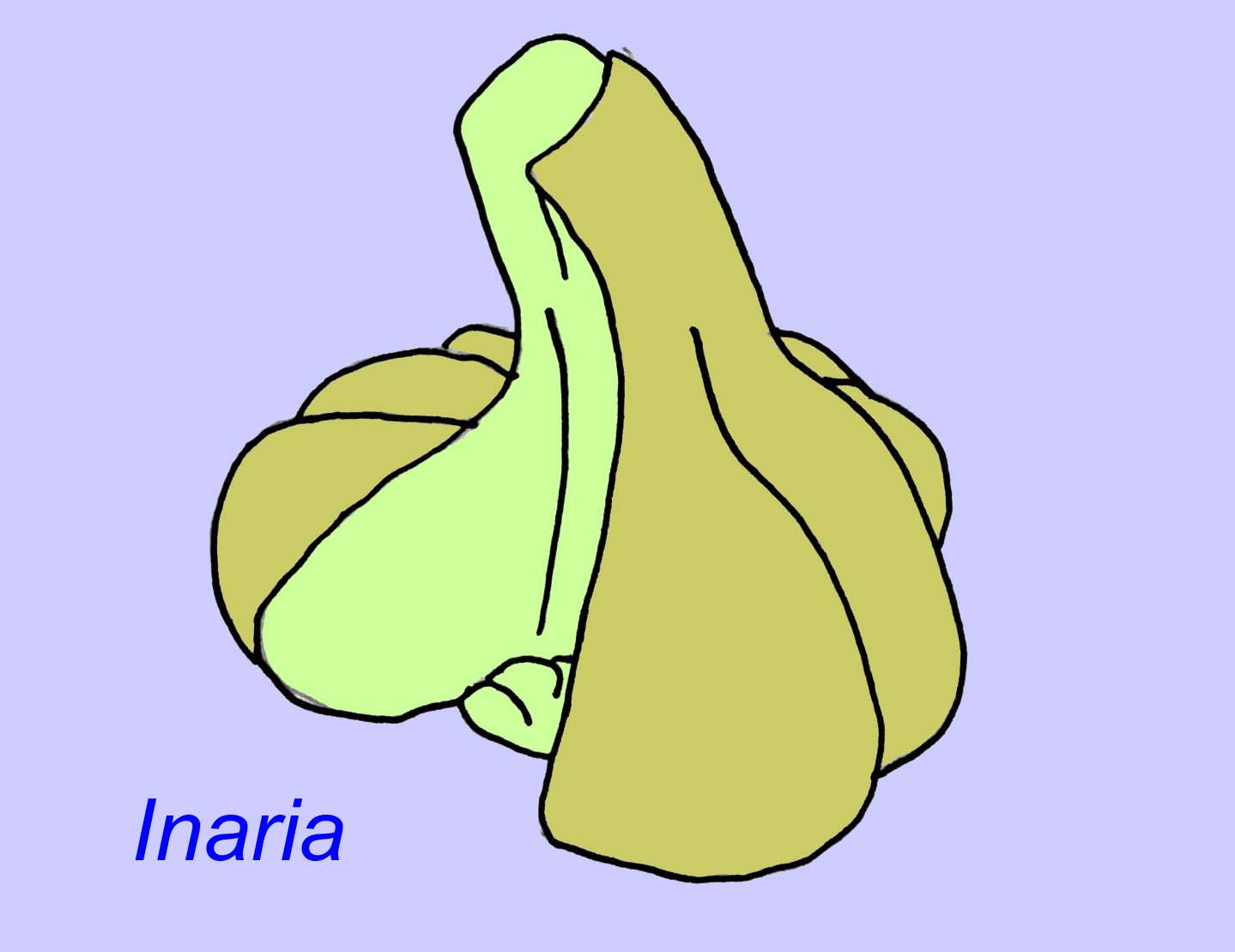
Mackenzia wydaje się też być knidarianinem (nawet jeśli nie, to prawie na pewno jest zwierzęciem metazoanicznym). Istnieją pośrednie dowody na obecność bolusów pokarmowych w jamie pokarmowej i muskulatury (Morris, 1993a). Sinocyclocyclicus może być bazalnym knidarianinem (lub nawet bazalnym metazoanem), którego pączkowanie wierzchołkowe jest podobne do tego u niektórych knidarian (np. koralowców tabularnych) (Xiao, 2000). Inaria to knidarianin z klasy Anthozoa z obrzeżem gębowym, gardzielą i komorą, w której mogły się znajdować organizmy fotosyntetyzujące, jak u wielu współczesnych koralowców (McMenaman, 1990, Gehling, 1988).
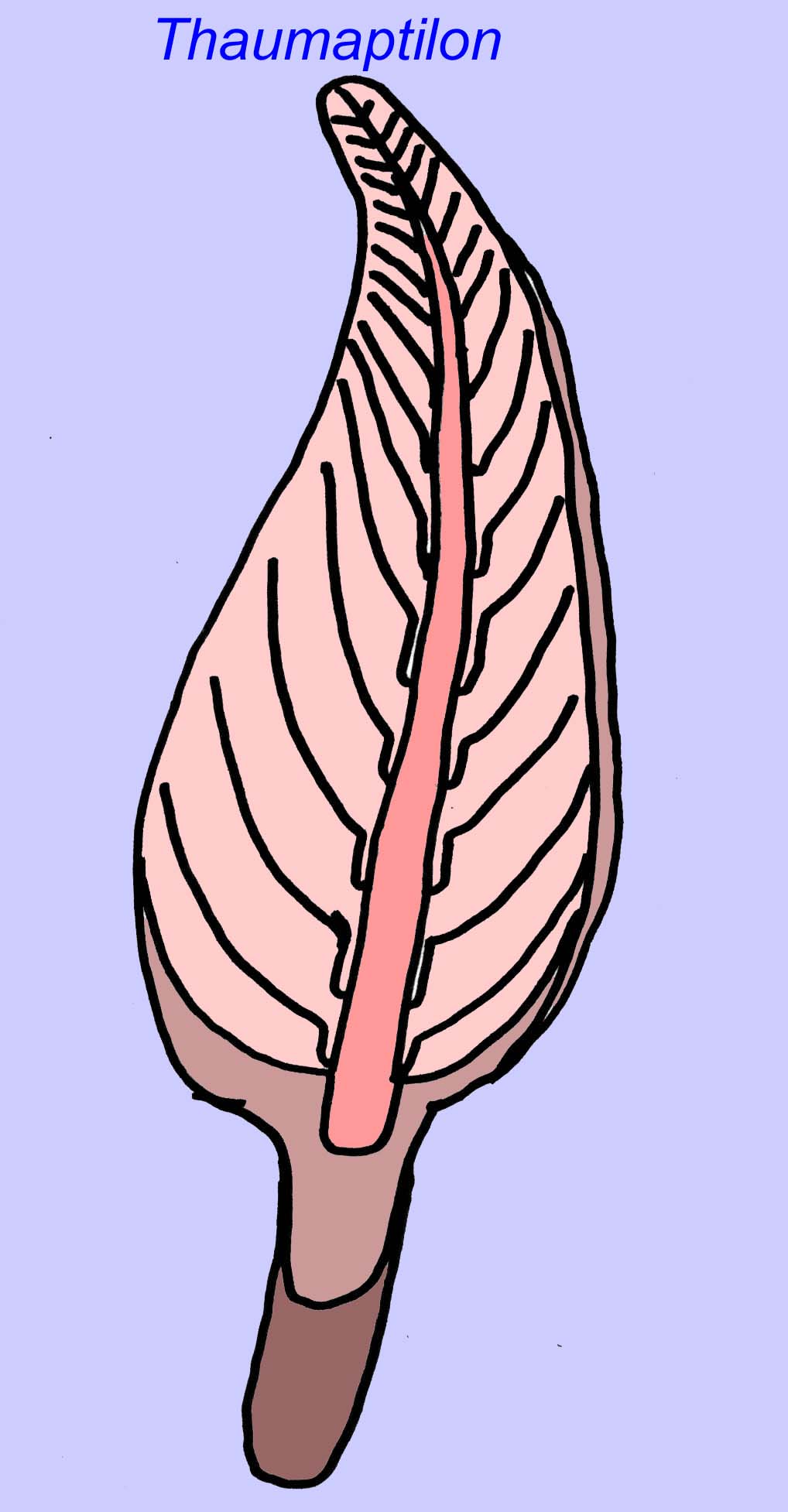
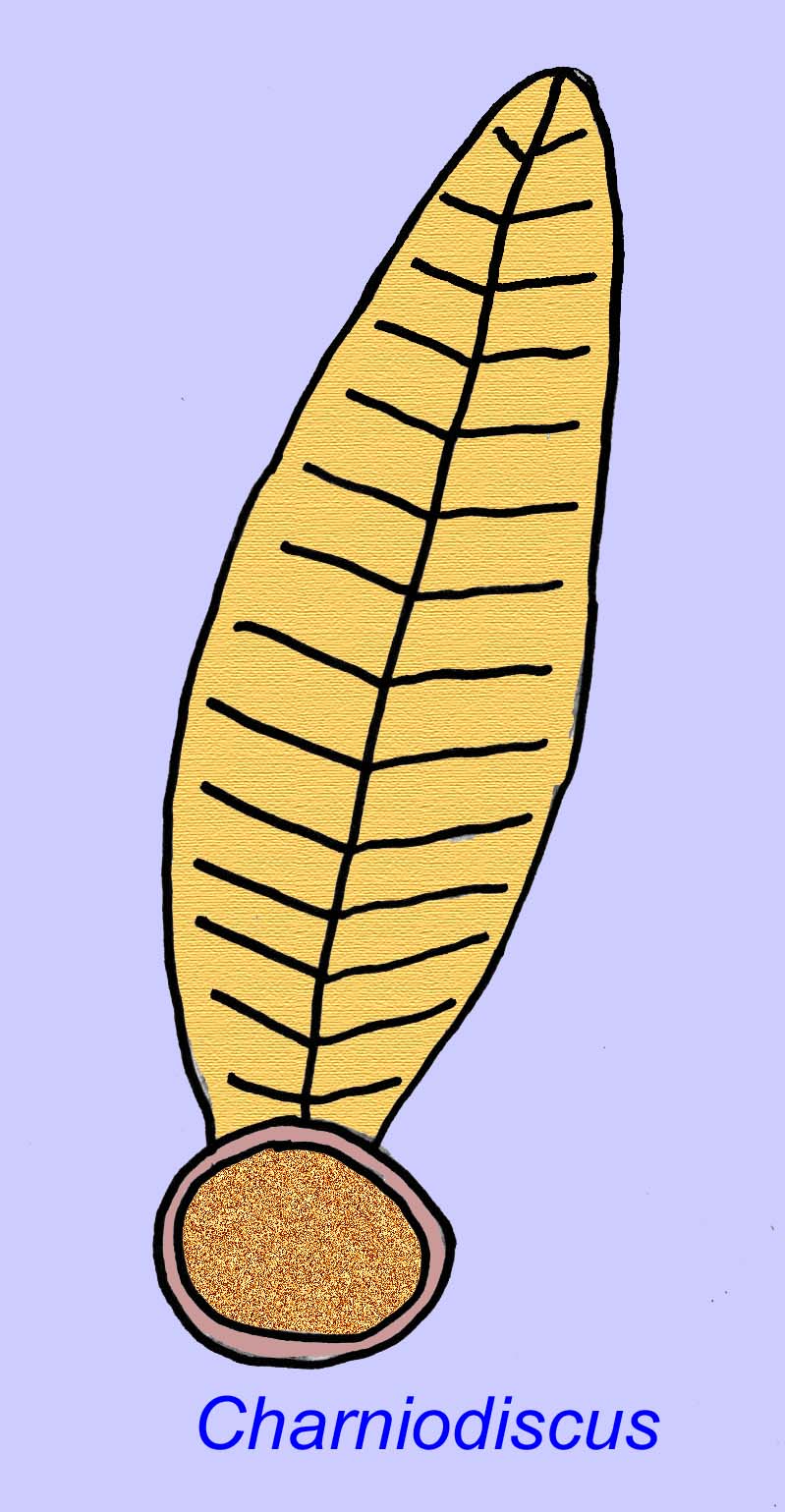
Thaumaptilon był przednio-podobnym zwierzęciem, które wydaje się spokrewnione ze współczesnymi piórami morskimi. Żywiłby się cząsteczkami i używałby holdfastu do mocowania się do dna (Morris, 1993a; 1993b). Czoło Charniodiscus mogło osiągać długość 1,2 metra (Sokolov, 1990).
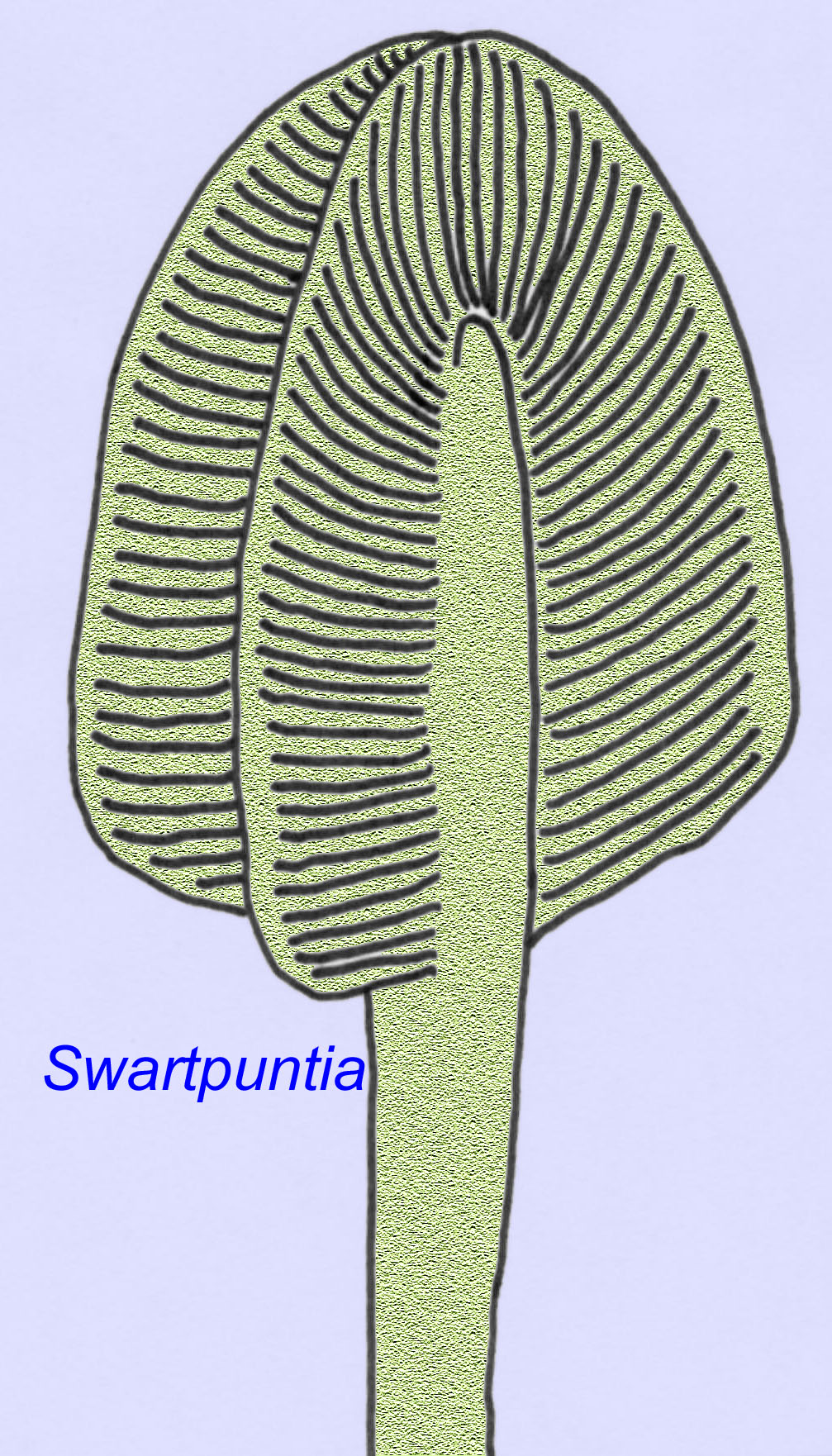
Wiele ediakarańskich zwierząt (takich jak Pteridinium, Swartpuntia, Charnia i Charniodiscus) posiada wyprostowaną łodygę połączoną z segmentowanymi płatkami.
Jeden okaz Charnodiscus mierzył 1,2 metra długości.
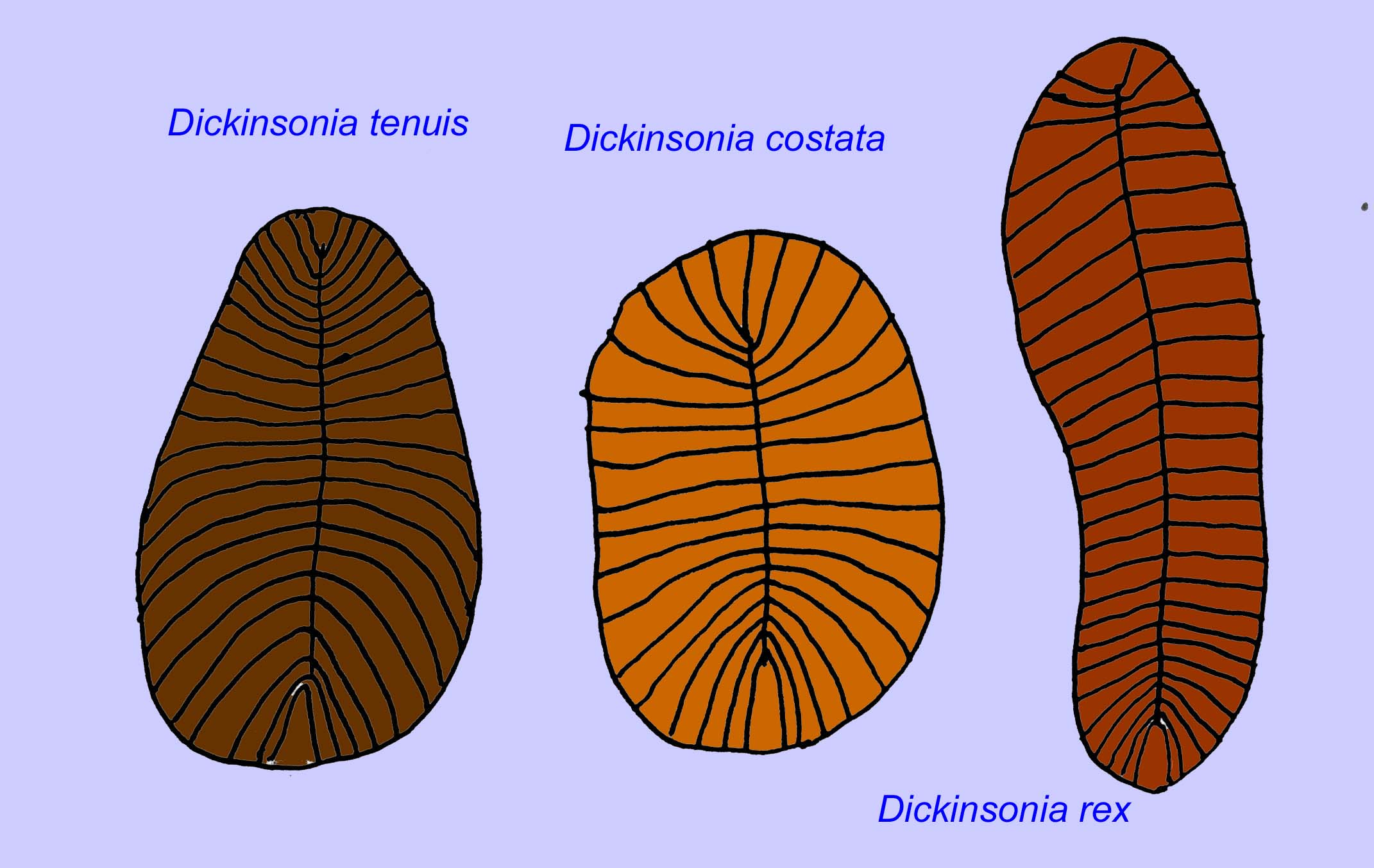
Anatomia mikroskopowa Swartpuntia jest bardzo podobna do anatomii Dickinsonia, wczesnego robaka (Narbonne, 1997; McCall, 2006). Skamieniałe embriony, które wydają się należeć do zwierząt z rzędu bilateralnych, znane są sprzed 570 milionów lat (Xiao, 1998).

WORKI
Ediakarańskie robaki znane są z Azji, Europy, Afryki, Australii i Ameryki Północnej (McCall, 2006). Wiele ediakarskich zwierząt było rurkowatych, takich jak Archaeichnium, Cloudina, Corumbella i Onuphionella. Onuphionella została porównana do robaków annelidów, natomiast Corumbella do rurek zbudowanych przez polipy niektórych knidarów. Podczas gdy Archaeichnium było elastyczne, Cloudina pokryta była zmineralizowanym szkieletem. Niektóre rurki Cloudina (maksymalna długość poniżej 4 cm) były annulowane, a inne gładkie, choć może to odzwierciedlać różnice w stanie zachowania (Hagadorn, 2000). Cloudina to pierwsze znane zwierzę ze zmineralizowanym szkieletem, a 3% okazów ma wywiercone otwory wskazujące na drapieżnictwo. Być może to właśnie drapieżnictwo przyczyniło się do powstania twardszych struktur typowych dla tak wielu zwierząt kambryjskich (Bengston, 1992). Cloudina znana jest z prekambru Ameryki Południowej, Europy, Bliskiego Wschodu i południowo-wschodniej Azji, a jej krewniacy znani są z wczesnego kambru (przed rozprzestrzenieniem się trylobitów) w Ameryce Północnej (Grant, 1990). Skamieniałości Cloudina znane są z okresu co najmniej 6 milionów lat przed początkiem kambru (Condon, 2005).


Oprócz dwóch uznanych gatunków Cloudina, szereg innych kopalnych gatunków posiadało zwapnione muszle w późnym prekambrze (McCall, 2006). Cloudina, Namacalanthus i Conotubus to szkieletowe zwierzęta prekambryjskie, które znane są także z wczesnego kambru (Shu, 2008).
Liczba robaków, takich jak Paleolina evenkiana i Planolites beverlyensis, została znaleziona pod koniec proterozoiku i w okresie kambryjskim. Skamieniałości planolitów mają szerokość 1-2 mm i długość od 5 do 13 mm (Bartley, 1998). Niesegmentowane skamieniałości, takie jak Protechirus, Platypholina i Vladimissa nie są dobrze poznane. Niektóre ze skamieniałości są segmentowane, jak Dickinsonia, która jest klasyfikowana jako robak annelidalny. Najmniejsze okazy Dickinsonia mają 4 mm średnicy, podczas gdy największe mają 1 metr długości. Pomimo długości tych robaków, były one bardzo cienkie, poniżej 3 mm. Dowody sugerują, że ilość tlenu w atmosferze pod koniec prekambru wynosiła tylko około jednej dziesiątej jego obecnego stężenia, a ekstremalna cienkość (i tym samym duża powierzchnia) tych robaków mogła być przystosowaniem do dyfuzji tlenu. Robaki te były powszechne i szeroko rozpowszechnione. Żywiły się szczątkami na dnie oceanu, biorąc pod uwagę osad, który często wypełniał ich jelita (Runnegar, 1982). Prosty charakter Dickinsonia może wskazywać, że jest to raczej knidarianin niż bilateran (Brasier, 2008).
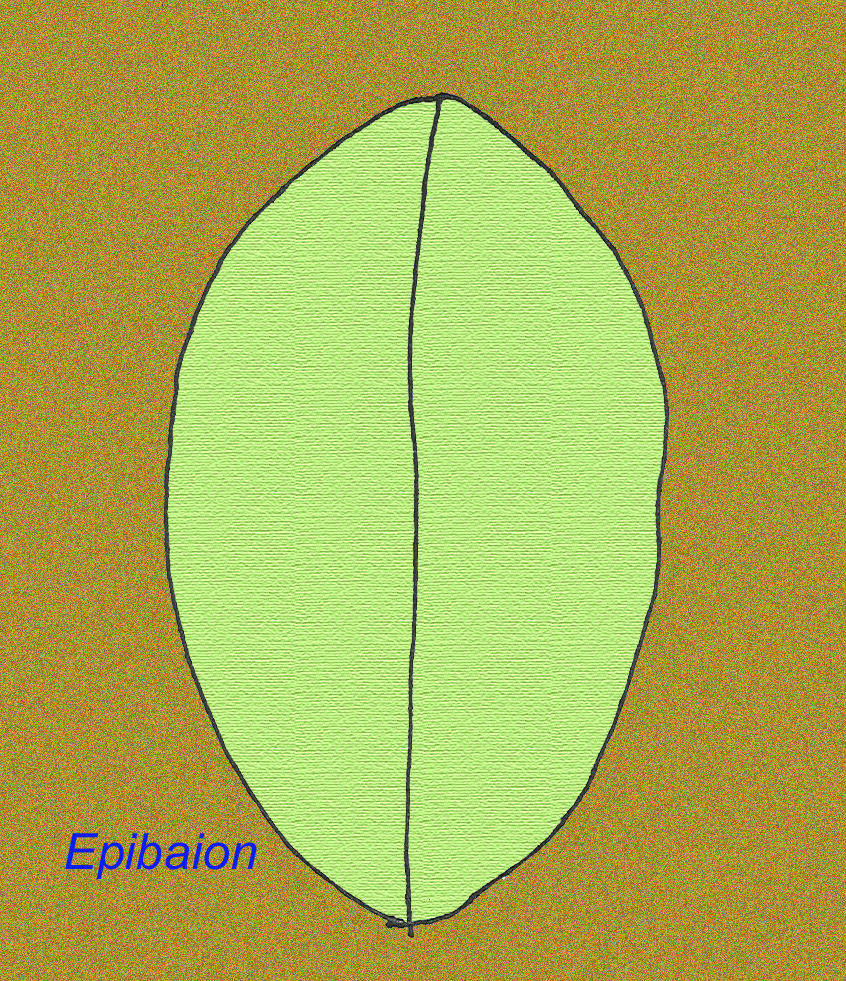
Duży osobnik z Ediacran o nazwie Epibaion axiferus pozostawił odciski o wymiarach do 4,3 m na .15 m (z drugim okazem o wymiarach 2,5 m na .35 m) (Ivantsov, 2002).


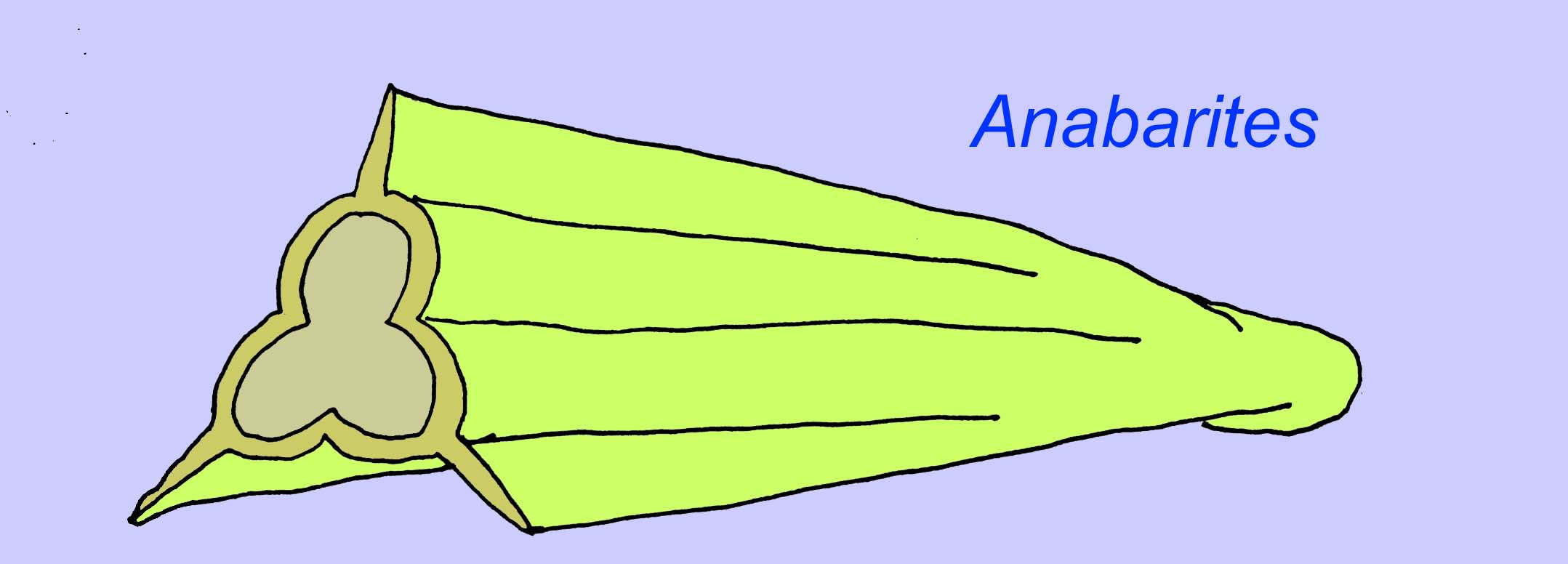
Anabaritydy to skamieniałości muszlowe z wczesnego kambru. Niektóre, takie jak anabaryty, mogą pochodzić z końca prekambru (McMenamin, 1990).
Śladowe skamieniałości nor robaków znaleziono w wielu miejscach proterozoicznych. Nory te zostały wykonane przez robaki bardziej zaawansowane niż płazińce. (Bartley, 1998, Narbonne, 1997, Hagadorn, 1998, McMenamin, 1996). Niektóre współczesne cnidaria mogą tworzyć proste nory i możliwe jest, że niektóre skamieniałości śladów nory zostały utworzone przez cnidaria, a nie bilaterany (Jensen, 2005). W niektórych norach robaków znajdują się odchody, co sugeruje istnienie układu pokarmowego podobnego do tego, który występuje u bardziej zaawansowanych robaków (Morris, 1993a; Valentine 1994). Skamieniałości embrionów robaków znaleziono już w najwcześniejszym kambrze (Bengston, 1997).
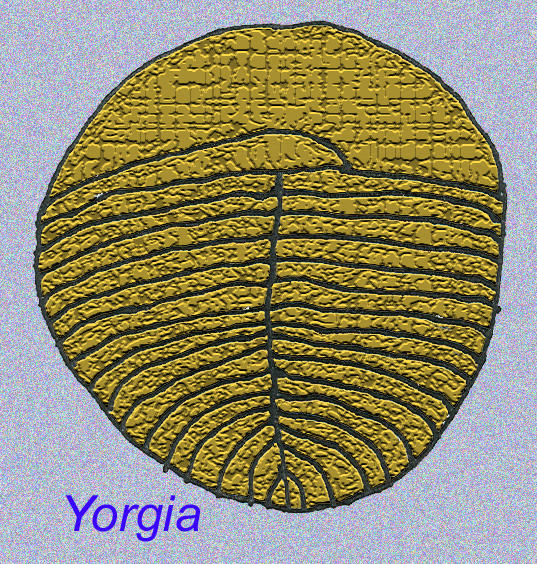
Czy wśród fauny EDIACARAN istniały zwierzęta bardziej zaawansowane niż robaki?
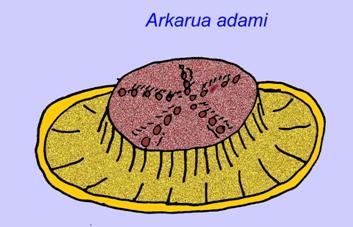
Niektóre analizy genetyczne sugerują, że rozdzielenie głównych grup zwierząt koelomaty nastąpiło przed kambrem (Gu, 1998). Wiele ediakarskich skamieniałości zostało zaklasyfikowanych do grup koelomatów, co potwierdza ten wniosek. Arkurua wykazuje podobieństwa do szkarłupni z 5-gałęzistą symetrią i podobieństwem do prymitywnych szkarłupni z kambru (edrioasteroidów). Być może te 5 struktur to ramiona żerne prowadzące do centralnego rowka (Gehling, 1987). Dwa rodzaje (Redkinia i Spriggina) mogą być bliskie podstawy linii stawonogów. Tribrachidium posiadający trzy wyrostki na jednej z powierzchni został odnaleziony w Australii i Europie (Wright, 1997b; McCall, 2006). Niektóre z ediakarskich faun miały plan ciała triradialny (Tribrachidium, Ventagyrus), tetraradialny (Conomedusies) i oktaradialny (Eoandromeda). Niektórzy interpretują je jako wymarłe linie cnidariów (Xiao, 2009).
Ediakarska skamieniałość Ausia została sklasyfikowana jako urochordate (Xiao, 2009).
Dwa gatunki, Phylozoon i Kimberella (zob. niżej), mogą być krewniakami segmentowanych robaków (annelidów) lub mięczaków. Kimberella ma symetrię dwustronną i możliwe jelito (Fedonkin, 1997). Wydaje się, że Kimberella była mięczakiem, który żył jako szybko pływający morski drapieżnik. Otwory wywiercone w muszlach Cloudina przypisano drapieżnikom z gatunku Edicaran (McCall, 2006). Skamieniałości embrionów z późnego prekambru tworzyły ciała biegunowe podobne do wielu mięczaków (Chen, 2006).
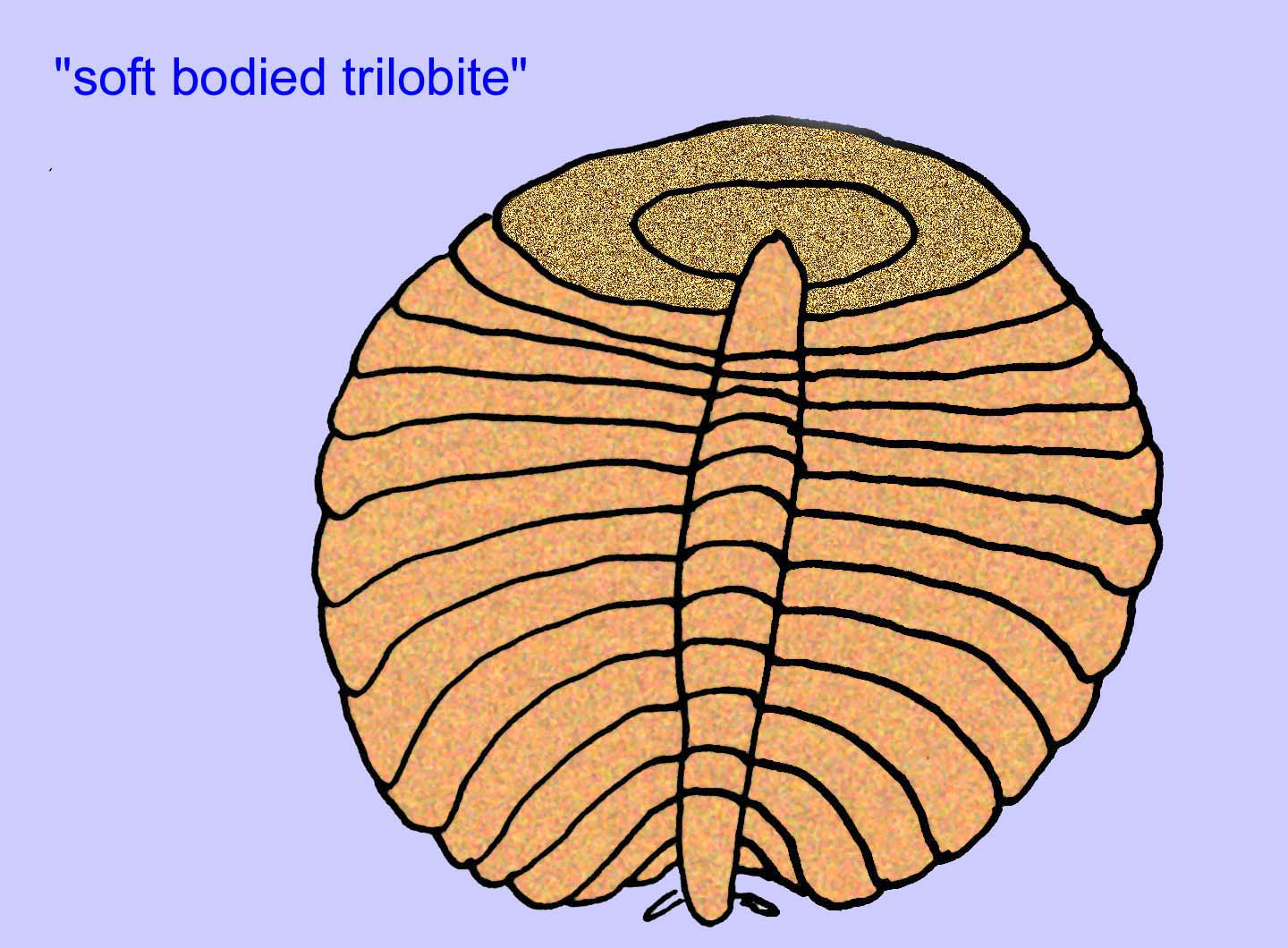
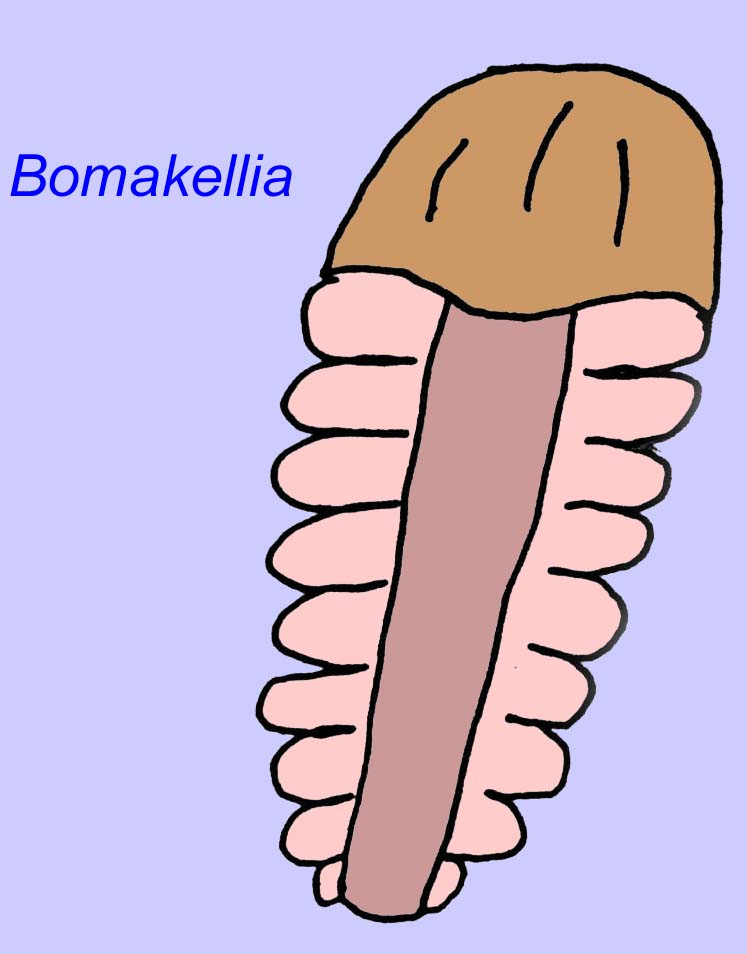
Inne skamieniałości mogą być związane ze stawonogami, z których niektóre (takie jak Bomakellia) mają strukturę porównywaną do tarczy głowy trylobitów (powszechny typ stawonogów ery paleozoicznej; McCall, 2006).
Niektóre z fauny ediakarskiej miały plan ciała trójpromienny (Tribrachidium, Ventagyrus), czteropromienny (Conomedusies) i ośmiopromienny (Eoandromeda). Niektórzy interpretują je jako wymarłe linie cnidariów (Xiao, 2009).